
| Hội chứng Smith–Lemli–Opitz | |
|---|---|
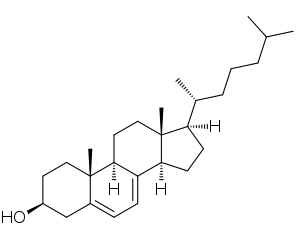 | |
| 7-Dehydrocholesterol | |
| Khoa/Ngành |
Di truyền học y khoa |
Hội chứng Smith–Lemli–Opitz (hay SLOS, khiếm khuyết 7-dehydrocholesterol reductase) là lỗi bẩm sinh của quá trình tổng hợp cholesterol. Đó là một hội chứng dị dạng di truyền lặn do một đột biến trong enzyme 7-Dehydrocholesterol reductase, hoặc DHCR7 . Nó gây ra một loạt các hiệu ứng, từ thiểu năng trí tuệ nhẹ và các vấn đề về hành vi đến dị tật gây chết người.
Dấu hiệu và triệu chứng
SLOS có thể biểu hiện khác nhau trong các trường hợp khác nhau, tùy thuộc vào mức độ nghiêm trọng của đột biến và các yếu tố khác. Ban đầu, bệnh nhân SLOS được phân thành hai loại (cổ điển và nặng) dựa trên các hành vi bên ngoài, đặc điểm thể chất và các đặc điểm lâm sàng khác. Kể từ khi phát hiện ra khiếm khuyết sinh hóa cụ thể chịu trách nhiệm về SLOS, bệnh nhân được đánh giá điểm nghiêm trọng dựa trên mức độ khuyết tật não, mắt, miệng và bộ phận sinh dục. Sau đó, nó được sử dụng để phân loại bệnh nhân bị SLOS nhẹ, cổ điển hoặc nặng.
Đặc điểm cơ thể
Các đặc điểm khuôn mặt phổ biến nhất của SLOS bao gồm microcephaly, thu hẹp bitem (giảm khoảng cách giữa các thái dương), ptosis, mũi ngắn và hếch, micrognathia, nếp gấp epicanthal và u mao mạch của mũi. Các đặc điểm khác bao gồm:
- tai thấp và xoay sau
- vòm miệng cao, hẹp, cứng
- sứt môi / vòm miệng
- agenesis hoặc hypoplasia của corpus callosum
- tiểu não hypoplasia
- tăng kích thước tâm thất
- giảm kích thước thùy trán
- tật nhiều ngón tay hoặc chân
- ngón tay cái ngắn, gần như được đặt
- dị tật ngón tay khác
- tật dính ngón của ngón chân thứ hai và thứ ba
- bộ phận sinh dục nam mơ hồ hoặc giống phụ nữ
- khuyết tật tim bẩm sinh
- Bất thường về thận, phổi, gan và mắt
Đặc điểm hành vi
Một số hành vi và thuộc tính nhất định thường thấy ở những bệnh nhân bị SLOS. Họ có thể có trí thông minh bình thường thấp, và phản ứng tiêu cực hoặc quá mẫn cảm với các kích thích giác quan khác nhau. Điều này đặc biệt đúng đối với một số kích thích thính giác và thị giác. Nhiều bệnh nhân thể hiện sự hung hăng và các hành vi tự gây thương tích và rối loạn giấc ngủ là phổ biến. Các hành vi cụ thể tương tự như những người mắc tự kỷ thường có mặt cũng như hiếu động, cung cấp hiểu biết di truyền và sinh học về rối loạn phổ tự kỷ. Các hành vi tự kỷ đặc trưng nhất của bệnh nhân SLOS là opisthokinesis (một chuyển động trên cơ thể), kéo dài của phần trên cơ thể và vẩy tay. Bệnh tự kỷ thường được chẩn đoán tách biệt với SLOS bằng cách sử dụng DSM-V và khoảng 50 mật75% bệnh nhân SLOS đáp ứng các tiêu chí cho bệnh tự kỷ.
Các hành vi khác liên quan đến SLOS có thể được liên kết trực tiếp với các bất thường về thể chất. Ví dụ, trẻ sơ sinh thường có vấn đề về ăn uống hoặc không dung nạp thức ăn, và bệnh nhân có thể yêu cầu tăng lượng calo do quá trình trao đổi chất một cách tăng tốc. Nhiễm trùng tái phát, bao gồm nhiễm trùng tai và viêm phổi, cũng rất phổ biến.
Kiểu hình sinh hóa
Cho rằng SLOS được gây ra bởi một đột biến trong một enzyme liên quan đến tổng hợp cholesterol, các đặc tính sinh hóa có thể dự đoán được. Hầu hết bệnh nhân đã giảm mức cholesterol trong huyết tương (hypocholesterolemia). Tuy nhiên, khoảng 10% có thể cho thấy mức cholesterol bình thường, và giảm nồng độ cholesterol không chỉ là biểu hiện của SLOS. Mức tiền chất cholesterol tăng cũng phổ biến trong SLOS. Cụ thể, mức độ 7-dehydrocholesterol khá cao đối với SLOS.
Chuyển hóa và chức năng cholesterol
Chuyển hóa
Cholesterol có thể có được thông qua chế độ ăn kiêng, nhưng nó cũng có thể được hình thành do sự trao đổi chất trong cơ thể. Chuyển hóa cholesterol chủ yếu diễn ra ở gan, với lượng đáng kể trong ruột là tốt. Cũng cần lưu ý rằng cholesterol không thể vượt qua hàng rào máu não, do đó trong não, sinh tổng hợp là nguồn cholesterol duy nhất.

Ở người, quá trình tổng hợp cholesterol bắt đầu bằng con đường mevalonate (xem sơ đồ), dẫn đến tổng hợp farnesyl pyrophosphate (FPP). Con đường này sử dụng hai acetyl-CoA và hai NADPH để tạo mevalonate, được chuyển hóa thành isopentenyl pyrophosphate (ba) ATP. Từ đó, cần có ba IPP để tạo một FPP. Sự kết hợp của hai FPP dẫn đến sự hình thành squalene; Điều này thể hiện bước cam kết đầu tiên đối với quá trình sinh tổng hợp cholesterol. Squalene dẫn đến việc tạo ra lanosterol, từ đó có nhiều con đường dẫn đến sinh tổng hợp cholesterol. Bước giới hạn tốc độ tổng hợp cholesterol là chuyển đổi 3-hydroxy-3-methylglutaryl-CoA (HMG-CoA) thành mevalonate, đây là bước đầu tiên trong con đường mevalonate được xúc tác bởi HMG -CoA reductase.


Thông qua một loạt các phản ứng phức tạp, lanosterol dẫn đến sự hình thành zymosterol. Như thể hiện trong một sơ đồ bên phải, tại thời điểm này, con đường phân kỳ. Ở người, con đường chính dẫn đến cholesterol được gọi là con đường Kandutsch-Russell. Zymosterol được chuyển hóa thành 5α-cholesta-7,24-dien-3β-ol, sau đó thành lathosterol, và sau đó thành 7-dehydrocholesterol hoặc 7-DHC. 7-DHC là tiền chất trực tiếp của cholesterol và enzyme DHCR7 chịu trách nhiệm chuyển 7-DHC thành cholesterol. DHCR7 làm giảm liên kết đôi trên carbon 7 của 7-DHC, dẫn đến sản phẩm chưa được ester hóa. Đột biến trong enzyme này chịu trách nhiệm cho một loạt các khiếm khuyết có trong SLOS. Trong một con đường khác dẫn đến tổng hợp cholesterol, DHCR7 là cần thiết để giảm 7-Dehydrodes ultraerol thành desimumerol.
Điều hòa
Sự điều hòa tổng hợp cholesterol rất phức tạp và xảy ra chủ yếu thông qua enzyme HMG-CoA reductase (chất xúc tác của bước giới hạn tốc độ). Nó liên quan đến một vòng feedback nhạy cảm với mức cholesterol trong tế bào. Bốn bước chính của điều hòa là:
- Sự tổng hợp của enzyme HMG-CoA reductase được kiểm soát bởi protein liên kết với yếu tố điều hòa sterol (SREBP). Đây là yếu tố phiên mã không hoạt động khi mức cholesterol cao và hoạt động khi mức cholesterol thấp. Khi mức cholesterol giảm, SREBP được giải phóng từ màng nhân hoặc lưới nội chất, sau đó nó di chuyển đến nhân và gây ra phiên mã của gen reductase HMG-CoA.
- Dịch mã (tạo enzyme từ bản phiên mã mRNA) của HMG-CoA reductase bị ức chế bởi các dẫn xuất của mevalonate và bởi cholesterol trong chế độ ăn uống.
- Sự giáng hóa của HMG-CoA reductase được kiểm soát chặt chẽ. Một phần của enzyme liên kết với mạng lưới nội chất cảm nhận các tín hiệu, chẳng hạn như mức cholesterol tăng, dẫn đến sự xuống cấp của nó hoặc proteolysis.
- Khi HMG-CoA reductase là phosphoryl hóa, hoạt động của nó giảm. Điều này có nghĩa là sự tổng hợp cholesterol bị giảm khi mức năng lượng tế bào (ATP) thấp.
Chức năng
Cholesterol là một lipid quan trọng liên quan đến chuyển hóa, chức năng tế bào và cấu trúc. Nó là một thành phần cấu trúc của màng tế bào, sao cho nó cung cấp cấu trúc và điều chỉnh tính lưu động của phospholipid hai lớp. Hơn nữa, cholesterol là thành phần trong lipid bè. Đây là các tập hợp của protein và lipid (bao gồm cả sphingolipid và cholesterol) trôi nổi trong màng tế bào và đóng vai trò trong việc điều chỉnh chức năng của màng. Các bè lipid được đặt hàng nhiều hơn hoặc cứng hơn so với màng đôi bao quanh chúng. Sự tham gia của họ vào quy định bắt nguồn chủ yếu từ sự liên kết của họ với protein; khi chất nền liên kết, một số protein có ái lực cao hơn để gắn vào bè lipid. Điều này mang lại cho chúng gần với các protein khác, cho phép chúng ảnh hưởng đến đường dẫn tín hiệu. Cholesterol đặc biệt hoạt động như một miếng đệm và keo cho bè lipid; thiếu cholesterol dẫn đến sự phân ly protein.
Do sự phổ biến của nó trong màng tế bào, cholesterol có liên quan nhiều đến các quá trình vận chuyển nhất định. Nó có thể ảnh hưởng đến chức năng của kênh ion và các chất vận chuyển màng khác. Ví dụ, cholesterol là cần thiết cho ligand hoạt động liên kết của thụ thể serotonin. Ngoài ra, nó có vẻ rất quan trọng trong exocytosis. Cholesterol điều chỉnh các thuộc tính của màng (như độ cong của màng) và có thể điều chỉnh sự hợp nhất của túi với màng tế bào. Nó cũng có thể tạo điều kiện cho việc tuyển dụng các phức hợp cần thiết cho exocytosis. Cho rằng nơ-ron phụ thuộc rất nhiều vào quá trình exocytosis để truyền xung động, cholesterol là một phần rất quan trọng của hệ thần kinh.

Một con đường đặc biệt có liên quan trong đó cholesterol diễn ra là Con đường báo hiệu Hedgekey. Con đường này rất quan trọng trong quá trình phát triển phôi và tham gia vào việc quyết định số phận của các tế bào (tức là, mô mà chúng cần di chuyển đến). Các protein của nhím cũng tham gia vào quá trình phiên mã các gen điều hòa tế bào tăng sinh và biệt hóa. Cholesterol rất quan trọng đối với con đường này bởi vì nó trải qua liên kết cộng hóa trị với protein Hedgehog, dẫn đến việc kích hoạt chúng. Không có cholesterol, hoạt động báo hiệu bị gián đoạn và sự biệt hóa tế bào có thể bị suy yếu.
Cholesterol là tiền chất cho nhiều phân tử quan trọng. Chúng bao gồm axit mật (quan trọng trong việc xử lý chất béo chế độ ăn uống), oxapseols, neurosteroids (liên quan đến dẫn truyền thần kinh và kích thích), glucocorticoid (liên quan đến miễn dịch và các quá trình viêm), mineralocorticoid (cân bằng thẩm thấu) và steroid sex (tức là estrogen và testosterone; phạm vi rộng của chức năng nhưng có liên quan đến sự phát triển bộ phận sinh dục). Cuối cùng, cholesterol là thành phần chính của myelin, một lớp bảo vệ xung quanh tế bào thần kinh. Quá trình myelin hóa xảy ra nhanh nhất trong quá trình phát triển trước khi sinh, có nghĩa là nhu cầu sinh tổng hợp cholesterol rất cao.
Đột biến DHCR7
DHCR7
Mã hóa gen DHCR7 (được dán nhãn là DHCR7 ) đã được sao chép vào năm 1998 và đã được ánh xạ tới nhiễm sắc thể 11q12-13. Nó dài 14100 cặp base DNA và chứa chín exon,mRNA tương ứng có chiều dài 2786 cặp base (trình tự DNA còn lại là nghiêm trọng). Cấu trúc của gen chuột DHCR7 rất giống với cấu trúc của gen người.
Các mức cao nhất của DHCR7 biểu hiện đã được phát hiện ở tuyến thượng thận, tinh hoàn, gan và trong mô não. Biểu hiện của nó được gây ra bởi giảm nồng độ sterol thông qua các protein liên kết điều hòa sterol (SREBP). Cũng có bằng chứng cho thấy hoạt động của nó có thể được điều chỉnh bằng phiên mã cụ thể của mô và ghép nối thay thế.
Như đã nêu ở trên, enzyme DHCR7 xúc tác cho việc giảm 7DHC thành cholesterol, cũng như giảm 7-dehydrodes ultraerol thành desimumerol. Nó yêu cầu NADPH làm đồng sáng lập cho việc giảm này và có thể liên quan đến hoạt động của cytochrom-P450 oxyoreductase. Nó cũng được cho là có chứa sắt. DHCR7 là một protein màng nguyên phân của mạng lưới nội chất và các mô hình máy tính đã dự đoán tới chín miền xuyên màng. DHCR7 hiệu quả nhất trong việc giảm 7DHC, nhưng được biết là làm giảm liên kết đôi carbon 7 của các sterol khác, cho thấy phạm vi cơ chất độ đặc hiệu. Phiên bản người của enzyme này được dự đoán có trọng lượng phân tử là 54.489 kDa và điểm đẳng điện là 9,05.
Trình tự amino acid mã hóa DHCR7 được dự đoán có chứa 475 amino acid, cũng như một số họa tiết protein. Nó chứa nhiều họa tiết reductase sterol, như mong đợi với chức năng của nó. Nó chứa một miền cảm ứng sterol tiềm năng (SSD), không rõ chức năng nhưng được cho là cần thiết để liên kết các chất nền sterol. Nó cũng bao gồm nhiều vị trí phosphoryl hóa, bao gồm các vị trí tiềm năng protein kinase C và tyrosine kinase (enzyme điều hòa chịu trách nhiệm cho quá trình phosphoryl hóa). Chức năng chính xác của phosphorylating DHCR7 vẫn chưa được biết, nhưng nó được cho là có liên quan đến việc điều chỉnh hoạt động của nó.
Đột biến và tỷ lệ mắc

Cho rằng SLOS là một rối loạn thể lặn, đột biến trong DHCR7 trên cả hai bản sao của nhiễm sắc thể 11 là cần thiết để bị rối loạn. Hơn 130 loại đột biến khác nhau đã được xác định.Missense đột biến (thay đổi nucleotide đơn dẫn đến mã cho một amino acid khác nhau) là phổ biến nhất, chiếm 87,6% phổ SLOS. Chúng thường làm giảm chức năng của enzyme nhưng có thể không ức chế hoàn toàn. Phần lớn phụ thuộc vào bản chất của đột biến (nghĩa là amino acid nào được thay thế và ở đâu). Đột biến Null ít phổ biến hơn nhiều, những đột biến này tạo ra một loại enzyme hoàn toàn rối loạn hoặc không có enzyme nào cả. Do đó, đột biến Missense có thể phổ biến hơn vì chúng ít gây chết người hơn đột biến vô nghĩa; đột biến vô nghĩa có thể đơn giản dẫn đến sảy thai tự nhiên.
IVS8-1G> C là đột biến được báo cáo thường xuyên nhất trong DHCR7 . Điều này phá vỡ sự tham gia của exon tám và chín, và dẫn đến việc chèn 134 nucleotide vào bảng điểm DHCR7 . Đây là một đột biến vô nghĩa, do đó, những bệnh nhân homozygous cho alen này bị ảnh hưởng nghiêm trọng. Người ta cho rằng đột biến này xảy ra lần đầu tiên trong Quần đảo Anh và nó có người mang gen (những người dị hợp tử cho tần số alen nhưng không bị ảnh hưởng) % cho người da trắng di sản châu Âu. Tần số đột biến khác nhau đối với các dân tộc khác nhau, tùy thuộc vào nguồn gốc của đột biến. Trong tất cả các quần thể da trắng, đột biến đặc biệt này có tần số sóng mang ước tính là 3%.
Đột biến phổ biến tiếp theo là 278C> T và dẫn đến threonine ở vị trí amino acid 93. Đây là đột biến Missense và có xu hướng liên quan đến các triệu chứng ít nghiêm trọng hơn. Đột biến này là đột biến thường gặp nhất ở bệnh nhân gốc Ý, Cuba và Địa Trung Hải.
Đột biến phổ biến thứ ba là 452G> A. Đột biến vô nghĩa này gây ra sự chấm dứt protein, do đó enzyme DHCR7 sẽ không được hình thành. Nó được cho là đã phát sinh ở Nam Ba Lan và phổ biến nhất ở Bắc Âu.
Các đột biến khác ít phổ biến hơn, mặc dù dường như nhắm mục tiêu vào các miền protein nhất định hơn so với các miền khác. Ví dụ, các họa tiết khử khử sterol là các vị trí phổ biến của đột biến. Nhìn chung, có tần số sóng mang ước tính (đối với bất kỳ đột biến DHCR7 gây SLOS) là 3-4% ở dân số da trắng (nó ít gặp hơn ở dân số châu Á và châu Phi ). Con số này cho thấy một ca sinh giả định tỷ lệ mắc trong khoảng từ 1/2500 đến 1/4500. Tuy nhiên, tỷ lệ mắc được đo là từ 1 / 10.000 đến 1 / 60.000 (nó khác nhau tùy thuộc vào di sản và gốc gác). Đây là thấp hơn nhiều so với dự kiến. Điều này cho thấy rằng nhiều trường hợp SLOS không bị phát hiện và có khả năng là do sảy thai tự nhiên do đột biến nghiêm trọng (sẩy thai) hoặc các trường hợp nhẹ không được chẩn đoán. Nữ giới thiếu các dị tật bộ phận sinh dục đặc trưng mà nam giới bị ảnh hưởng và do đó ít có khả năng được chẩn đoán chính xác.
Sinh bệnh học
Cho rằng chức năng của cholesterol bao gồm một phạm vi rất rộng, không có khả năng các triệu chứng của SLOS là do một cơ chế phân tử duy nhất. Một số tác dụng phân tử chưa được biết, nhưng có thể được ngoại suy dựa trên vai trò của cholesterol. Nói chung, các tác động tiêu cực là do mức cholesterol giảm và mức độ tiền chất cholesterol tăng lên - đáng chú ý nhất là 7DHC. Mặc dù 7DHC có cấu trúc tương tự như cholesterol và có khả năng đóng vai trò thay thế, nhưng tác dụng của việc này vẫn đang được nghiên cứu.
Hầu hết bệnh nhân bị SLOS biểu hiện giảm mức cholesterol, đặc biệt là trong não (nơi mức cholesterol chủ yếu dựa vào tổng hợp mới). Điều này cũng có nghĩa là bất kỳ dẫn xuất sterol nào của cholesterol cũng sẽ làm giảm nồng độ. Ví dụ, có thể thấy mức độ giảm neurosteroid trong SLOS. Đây là những lipid tham gia truyền tín hiệu trong não và phải được sản xuất trong chính não. Chúng chịu trách nhiệm tương tác với thụ thể steroid hạt nhân và liên kết với dẫn truyền thần kinh - các kênh ion bị kiểm soát. Cụ thể, họ điều chỉnh tác động của GABA và NMDA, dẫn đến hiệu ứng làm dịu, cải thiện trí nhớ và hơn thế nữa. Do đó, do một số đặc điểm của SLOS trái ngược với những tác động này (tăng động, lo lắng), việc giảm neurosteroid có thể ảnh hưởng đến cả sự phát triển và hành vi thần kinh.

Hơn nữa, như đã nêu ở trên, cholesterol là một khía cạnh quan trọng trong tín hiệu Hedgehog. Với mức cholesterol thấp hơn, protein nhím sẽ không trải qua quá trình điều chỉnh cộng hóa trị cần thiết và kích hoạt tiếp theo. Điều này sẽ dẫn đến sự phát triển phôi bị suy giảm, và có thể góp phần vào dị tật bẩm sinh quan sát được trong SLOS. Một protein báo hiệu con nhím đặc biệt, sonic hedgeoose (SHH), rất quan trọng trong mô hình của hệ thống thần kinh trung ương, đặc điểm khuôn mặt và chân tay. Các protein hedgehog khác có thể liên quan đến sự phát triển của đường sinh dục và xương.
Nồng độ sterol thay đổi trong SLOS đặc biệt liên quan đến màng tế bào, được tạo ra chủ yếu từ lipid. Bệnh nhân SLOS có thể thấy màng tế bào có đặc tính hoặc thành phần bất thường, và mức cholesterol giảm ảnh hưởng lớn đến sự ổn định và protein của bè lipid. Mặc dù có sự tương đồng về cấu trúc, 7DHC không thể thay thế cholesterol trong bè lipid. Ngoài ra, việc thiếu cholesterol góp phần làm tăng tính lưu động của màng tế bào và có thể gây ra sự tiết bất thường hạt. Tất cả những thay đổi này trong màng có thể góp phần thay đổi chức năng vận chuyển được quan sát trong SLOS. Chúng có thể gây ra khiếm khuyết trong IgE qua trung gian thụ thể tế bào mast và sản xuất cytokine, là những tế bào liên quan đến phản ứng dị ứng và miễn dịch. Thụ thể NMDA bị ảnh hưởng, cũng như khả năng liên kết của hippocampal thụ thể serotonin.Tương tác giữa tế bào với tế bào, rất quan trọng trong quá trình phát triển, có thể bị suy yếu.Exocytosis trong synap vesicle đã được chứng minh là đã giảm, có khả năng do phản ứng tổng hợp túi tinh bị suy yếu đến màng tế bào hoặc tái chế mụn nước kém. Cuối cùng, cholesterol rất phổ biến trong myelin, do đó, bệnh nhân SLOS cho thấy giảm quá trình myel hóa bán cầu não, dây thần kinh ngoại biên và dây thần kinh sọ.
Ngoài việc giảm mức cholesterol, nhiều triệu chứng trong SLOS xuất phát từ tác dụng độc hại của 7DHC. 7DHC được biết là làm suy yếu sự vận chuyển cholesterol nội bào. Nó cũng làm tăng sự thoái hóa của HMG-CoA reductase (enzyme xúc tác cho bước giới hạn tốc độ trong quá trình tổng hợp cholesterol). 7DHC dẫn đến các dẫn xuất mới oxapseol và steroid, và nhiều chức năng hoặc tác dụng của chúng chưa được biết đến. Một phát hiện rất quan trọng đối với 7DHC là nó là loại lipid dễ phản ứng nhất đối với peroxid hóa lipid và dẫn đến hệ thống stress oxy hóa. Sự peroxy hóa lipid được biết là phá hủy màng của cả tế bào và màng organelle. Đạo hàm của 7DHC được sử dụng để chỉ ra stress oxy hóa là 3β, 5α-dihydroxy-cholest-7-en-6-one (DHCEO); nó được hình thành từ một sản phẩm chính của peroxid hóa 7DHC, 7-DHC-5α, 6α-epoxide. DHCEO độc với tế bào thần kinh vỏ não và tế bào thần kinh đệm, và tăng tốc biệt hóa và arborization. Thông qua căng thẳng oxy hóa, 7DHC được cho là nguyên nhân làm tăng nhạy cảm ánh sáng thể hiện ở bệnh nhân SLOS. Tiếp xúc bình thường UVA có thể dẫn đến căng thẳng oxy hóa trong các tế bào da. Cho rằng 7DHC dễ bị oxy hóa hơn, nó làm tăng tác dụng của UVA, dẫn đến tăng quá trình oxy hóa lipid màng và tăng sản xuất loại oxy phản ứng (ROS).
Thông thường, sự thay đổi nhiều hơn mức 7DHC và cholesterol dẫn đến các triệu chứng SLOS nghiêm trọng hơn. Mức độ của các chất chuyển hóa này cũng tương ứng với mức độ nghiêm trọng của đột biến (vô nghĩa so với sai lầm); một số đột biến của DHCR7 vẫn có thể cho thấy sự tổng hợp cholesterol còn lại, và một số khác thì không. Tuy nhiên, ngay cả những cá nhân có cùng đột biến hoặc kiểu gen vẫn có thể cho thấy sự thay đổi trong các triệu chứng của họ. Điều này có thể là do các yếu tố của người mẹ, chẳng hạn như chuyển cholesterol cho thai nhi trong thai kỳ, cũng như lượng cholesterol có trong não trước khi hàng rào máu não hình thành trước khi sinh. Tốc độ tích lũy và bài tiết các chất chuyển hóa độc hại có thể khác nhau tùy theo từng người. Người mẹ apolipoprotein E cũng có liên quan đến sự biến đổi cá nhân trong SLOS, mặc dù bản chất chính xác của mối quan hệ này chưa được biết. Có nhiều yếu tố góp phần vào phổ hiệu ứng rộng trong SLOS chưa được phát hiện.
Sàng lọc và chẩn đoán
Trước sinh
Chỉ số sinh hóa đặc trưng nhất của SLOS là nồng độ tăng 7DHC (giảm cholesterol cũng rất điển hình, nhưng cũng xuất hiện trong các rối loạn khác). Do đó, prenatally, SLOS được chẩn đoán khi tìm thấy tỷ lệ 7DHC tăng cao: tổng tỷ lệ sterol trong các mô của thai nhi, hoặc tăng mức 7DHC trong nước ối. Tỷ lệ 7DHC: tổng sterol có thể được đo tại 11 đỉnh12 tuần thời gian mang thai bằng lấy mẫu lông nhung màng đệm, và 7DHC tăng trong nước ối có thể được đo trong 13 tuần. Hơn nữa, nếu biết được các đột biến của cha mẹ, xét nghiệm DNA của nước ối hoặc mẫu lông nhung màng đệm có thể được thực hiện.

Chọc ối (quá trình lấy mẫu nước ối) và lấy mẫu lông nhung màng đệm có thể được thực hiện cho đến khoảng 3 tháng sau khi mang thai. Cho rằng SLOS là một hội chứng rất nghiêm trọng, cha mẹ có thể muốn chọn bỏ thai nếu thai nhi bị ảnh hưởng. Lấy mẫu nước ối và lấy mẫu lông nhung màng đệm để lại rất ít thời gian để đưa ra quyết định này (việc phá thai trở nên khó khăn hơn khi thai tiến triển), và cũng có thể gây ra những rủi ro nghiêm trọng cho mẹ và bé. Vì vậy, có một mong muốn rất lớn đối với các xét nghiệm chẩn đoán trung bình không xâm lấn. Kiểm tra nồng độ sterol trong nước tiểu của mẹ là một cách tiềm năng để xác định SLOS trước khi sinh. Khi mang thai, thai nhi chỉ chịu trách nhiệm tổng hợp cholesterol cần thiết để sản xuất estriol. Một bào thai với SLOS không thể tạo ra cholesterol và có thể sử dụng 7DHC hoặc 8DHC làm tiền chất cho estriol thay thế. Điều này tạo ra 7 hoặc 8-dehydrosteroid (như 7-dehydroestriol), có thể xuất hiện trong nước tiểu của mẹ. Đây là những chất chuyển hóa mới do sự hiện diện của liên kết đôi thường giảm ở carbon 7 (gây ra bởi sự không hoạt động của DHCR7) và có thể được sử dụng làm chỉ số của SLOS. Các dẫn xuất cholesterol khác có liên kết đôi ở vị trí thứ 7 hoặc thứ 8 và có trong nước tiểu của mẹ cũng có thể là chỉ số của SLOS. 7- và 8-dehydropregnanetriols đã được chứng minh là có trong nước tiểu của những bà mẹ có thai nhi bị ảnh hưởng nhưng không phải là thai nhi không bị ảnh hưởng, và do đó được sử dụng trong chẩn đoán. Những mergadiene này bắt nguồn từ bào thai và đi qua nhau thai trước khi đến tay người mẹ. Sự bài tiết của chúng chỉ ra rằng cả nhau thai và các cơ quan của mẹ đều không có các enzyme cần thiết để giảm liên kết đôi của các chất chuyển hóa mới này.
Sau sinh
Nếu SLOS không bị phát hiện cho đến sau khi sinh, chẩn đoán có thể dựa trên các đặc điểm vật lý đặc trưng cũng như tìm thấy mức độ huyết tương tăng 7DHC.
Có nhiều cách khác nhau để phát hiện nồng độ 7DHC trong huyết tương, một cách là sử dụng thuốc thử Liebermann-Burchard (LB). Đây là một xét nghiệm đo màu đơn giản được phát triển với mục đích sử dụng để sàng lọc quy mô lớn. Khi được xử lý bằng thuốc thử LB, các mẫu SLOS chuyển sang màu hồng ngay lập tức và dần dần chuyển sang màu xanh; mẫu máu bình thường ban đầu không màu và phát triển màu xanh nhạt. Mặc dù phương pháp này có những hạn chế và không được sử dụng để đưa ra chẩn đoán xác định, nhưng nó có sức hấp dẫn ở chỗ nó là phương pháp nhanh hơn nhiều so với sử dụng nuôi cấy tế bào.
Một cách khác để phát hiện 7DHC là thông qua sắc ký khí, một kỹ thuật được sử dụng để tách và phân tích các hợp chất. Phương pháp sắc ký khí / khối phổ theo dõi ion được chọn (SIM-GC / MS) là phiên bản rất nhạy của sắc ký khí và cho phép phát hiện các trường hợp SLOS nhẹ. Các phương pháp khác bao gồm quang phổ khối lượng thời gian bay, chùm hạt LC / MS, ion hóa electrospray tandem MS và tia cực tím có thể nhìn thấy bằng tia cực tím , tất cả trong số đó có thể được sử dụng trên cả mẫu máu, nước ối hoặc lông nhung màng đệm. Đo nồng độ axit mật trong nước tiểu bệnh nhân, hoặc nghiên cứu hoạt động DCHR7 trong nuôi cấy mô cũng là những kỹ thuật chẩn đoán sau sinh phổ biến.
Điều trị
Quản lý các cá nhân với SLOS rất phức tạp và thường cần một đội ngũ chuyên gia. Một số dị tật bẩm sinh (hở vòm miệng) có thể được sửa chữa bằng phẫu thuật. Các phương pháp điều trị khác vẫn chưa được chứng minh thành công trong các nghiên cứu ngẫu nhiên, tuy nhiên về mặt giai đoạn chúng dường như gây ra sự cải thiện.
Bổ sung cholesterol
Hiện nay, hình thức điều trị SLOS phổ biến nhất liên quan đến bổ sung cholesterol trong chế độ ăn uống. Các báo cáo giai thoại chỉ ra rằng điều này có một số lợi ích; nó có thể dẫn đến tăng trưởng, giảm khó chịu, cải thiện tính xã hội, ít hành vi tự gây tổn thương, ít phòng thủ xúc giác, ít nhiễm, nhiều cơ bắp hơn, ít nhạy cảm ánh sáng và ít hơn hành vi tự kỷ. Bổ sung cholesterol bắt đầu với liều 40 -50 mg / kg / ngày, tăng khi cần thiết. Nó được quản lý thông qua việc tiêu thụ thực phẩm có hàm lượng cholesterol cao (trứng, kem, gan) hoặc dưới dạng cholesterol thực phẩm tinh khiết. Trẻ nhỏ và trẻ sơ sinh có thể cần cho ăn bằng ống. Tuy nhiên, cholesterol trong chế độ ăn kiêng không làm giảm mức 7DHC, không thể vượt qua hàng rào máu não và dường như không cải thiện kết quả phát triển. Một nghiên cứu thực nghiệm cho thấy việc bổ sung cholesterol không cải thiện chậm phát triển, bất kể độ tuổi bắt đầu. Điều này có thể là do hầu hết sự chậm phát triển bắt nguồn từ dị tật của não, mà cholesterol trong chế độ ăn uống không thể cải thiện do không có khả năng vượt qua hàng rào máu não..

Liệu pháp simvastatin
Chất ức chế HMG-CoA reductase đã được kiểm tra để điều trị SLOS. Cho rằng xúc tác bước giới hạn tốc độ trong tổng hợp cholesterol, ức chế nó sẽ làm giảm sự tích tụ các chất chuyển hóa độc hại như 7DHC.Simvastatin là một chất ức chế được biết đến của HMG-CoA reductase, và quan trọng nhất là có thể vượt qua hàng rào máu não. Nó đã được báo cáo để giảm mức độ 7DHC, cũng như tăng mức độ cholesterol. Nồng độ cholesterol tăng là do ảnh hưởng của simvastatin đối với sự biểu hiện của các gen khác nhau. Simvastatin làm tăng biểu hiện của DHCR7 , có khả năng dẫn đến tăng hoạt động của DHCR7. Nó cũng đã được chứng minh là làm tăng sự biểu hiện của các gen khác liên quan đến tổng hợp và hấp thu cholesterol. Tuy nhiên, những lợi ích này phụ thuộc vào lượng tổng hợp cholesterol còn lại. Bởi vì một số cá nhân sở hữu các đột biến ít nghiêm trọng hơn và chứng minh một số lượng hoạt động DCHR7, những người này được hưởng lợi nhiều nhất từ liệu pháp simvastatin vì họ vẫn có một enzyme hoạt động một phần. Đối với các cá nhân không có hoạt động DCHR7 còn lại, chẳng hạn như homozygous đối với null alen hoặc đột biến, liệu pháp simvastatin thực sự có thể gây độc. Điều này nhấn mạnh tầm quan trọng của việc xác định kiểu gen cụ thể của bệnh nhân SLOS trước khi điều trị. Vẫn chưa biết liệu simvastatin sẽ cải thiện các khiếm khuyết về hành vi hoặc học tập trong SLOS.
Bổ sung chất chống oxy hóa
Chất chống oxy hóa là những chất ức chế quá trình oxy hóa của các phân tử hoặc làm giảm các chất chuyển hóa đã bị oxy hóa trước đó. Cho rằng một số triệu chứng của SLOS được cho là kết quả từ peroxidation lipid peroxidation của 7DHC và các dẫn xuất của nó, ức chế peroxid hóa này có thể có tác dụng có lợi. Chất chống oxy hóa đã được chứng minh là làm tăng mức độ của các bản sao lipid trong các tế bào SLOS, các bản sao này đóng vai trò trong sinh tổng hợp lipid (cholesterol) và được biết là được điều chỉnh xuống trong SLOS. Hơn nữa, vitamin E đặc biệt được biết là làm giảm nồng độ DHCEO, một chỉ số của stress oxy hóa trong SLOS, cũng như thể hiện những thay đổi có lợi trong biểu hiện gen. Vitamin E dường như là chất chống oxy hóa mạnh nhất để điều trị SLOS, và trong các mô hình chuột đã làm giảm mức oxapseol trong não. Tuy nhiên, chất chống oxy hóa chỉ được nghiên cứu trên mô hình động vật của SLOS hoặc tế bào SLOS bị cô lập. Do đó, ý nghĩa lâm sàng và tác dụng phụ tiêu cực của chúng vẫn chưa được biết và việc sử dụng chúng vẫn chưa được nghiên cứu ở người.
Cân nhắc thêm
Khi điều trị SLOS, một vấn đề tái phát là liệu sự thiếu hụt về trí tuệ và hành vi có phải là do các vấn đề phát triển cố định (nghĩa là dị tật não cố định) hoặc do nồng độ sterol bất thường liên tục làm gián đoạn chức năng bình thường của não và các mô khác. Nếu điều này là đúng, thì các phương pháp điều trị làm thay đổi nồng độ và tỷ lệ sterol, đặc biệt là trong não, có thể sẽ cải thiện kết quả phát triển của bệnh nhân. Tuy nhiên, nếu điều này là đúng, thì điều trị có khả năng chỉ giúp với các triệu chứng chứ không phải với các thiếu hụt phát triển cụ thể.
Mô hình / nghiên cứu động vật
Động vật phổ biến nhất được sử dụng để nghiên cứu SLOS là chuột. Theo BioCyc, quá trình sinh tổng hợp cholesterol ở chuột rất giống với con người. Quan trọng nhất, chuột có cả DHCR7 (enzyme chịu trách nhiệm về SLOS) và HMG-CoA reductase (bước giới hạn tốc độ tổng hợp cholesterol. Chuột cống tương tự như chuột và cũng đã được sử dụng. Có hai cách phổ biến trong đó mô hình động vật của SLOS được tạo ra. Đầu tiên là sử dụng teratogens, thứ hai là sử dụng các thao tác di truyền để tạo đột biến trong gen DHCR7 .
Mô hình gây quái thai
Các mô hình quái thai được tạo ra bằng cách cho chuột hoặc chuột mang thai ức chế của DCHR7. Hai chất ức chế phổ biến là BM15766 (4 - (2- [1- (4-chlorocinnamyl) piperazin-4-yl] ethyl) -benzoic acid) và AY9944 (trans-l, 4-bis (2-chlorobenzylaminomethy1) cyclohexane). Các hợp chất này có tính chất hóa học và vật lý khác nhau, nhưng gây ra tác dụng tương tự. AY9944 đã được chứng minh là gây ra holoprosencephaly và dị tật tình dục tương tự như những gì nhìn thấy ở người bị SLOS. Nó cũng được biết là gây ra sự suy yếu ở thụ thể serotonin, một khiếm khuyết khác thường thấy ở bệnh nhân SLOS. BM15766 đã tạo ra sự thiếu hụt cholesterol và axit mật được thấy ở những bệnh nhân SLOS có homozygous. Tất cả các mô hình gây quái thai có thể được sử dụng hiệu quả để nghiên cứu SLOS; tuy nhiên, chúng có mức 7-DHC và 8-DHC thấp hơn so với người ta thấy. Điều này có thể được giải thích bởi thực tế là con người trải qua một khối vĩnh viễn trong hoạt động DHCR7 của họ, nơi chuột và chuột được điều trị bằng chất ức chế chỉ trải qua các khối thoáng qua. Hơn nữa, các loài chuột và chuột khác nhau có nhiều kháng với quái thai, và có thể kém hiệu quả như các mô hình của SLOS. Các mô hình gây quái thai được sử dụng phổ biến nhất để nghiên cứu các tác động dài hạn hơn của SLOS, bởi vì chúng tồn tại lâu hơn các mô hình di truyền. Ví dụ, một nghiên cứu đã kiểm tra sự thoái hóa võng mạc của SLOS, ở chuột không xảy ra cho đến ít nhất một tháng sau khi sinh.
Mô hình di truyền
Mô hình di truyền của SLOS được tạo ra bởi gõ cửa gen DHCR7 . Một nghiên cứu đã sử dụng tái tổ hợp tương đồng để phá vỡ DCHR7 trên chuột tế bào gốc phôi. Tương tự như những gì được tìm thấy ở người, chuột dị hợp tử (chỉ có một alen đột biến) là bình thường. Mặc dù những chú chó con này đã chết trong ngày đầu tiên của cuộc đời do không thể cho ăn, chúng đã cho thấy những đặc điểm tương tự như con người với SLOS. Họ đã giảm mức cholesterol, tăng mức 7 và 8DHC, cho thấy sự tăng trưởng ít hơn và trọng lượng sơ sinh nhỏ hơn, có dị tật craniofacial và ít vận động hơn. Nhiều người cũng có sứt môi và giảm phản ứng thần kinh với glutamate. Tuy nhiên, nhìn chung, những con chó con có các đặc điểm dị hình ít hơn so với bệnh nhân mắc SLOS ở người; họ không có dị tật chân tay, thận, thượng thận hoặc hệ thần kinh trung ương. Điều này được giải thích bởi thực tế là ở loài gặm nhấm, cholesterol của mẹ có thể vượt qua nhau thai, và thực sự có vẻ rất cần thiết cho sự phát triển của thai nhi. Ở người, rất ít cholesterol của mẹ được chuyển đến thai nhi. Tóm lại, mô hình chuột di truyền rất hữu ích để giải thích sinh lý bệnh thần kinh của SLOS.
Khám phá
Nhiều khám phá trong nghiên cứu SLOS đã được thực hiện bằng cách sử dụng mô hình động vật. Chúng đã được sử dụng để nghiên cứu các kỹ thuật điều trị khác nhau, bao gồm hiệu quả của liệu pháp simvastatin. Các nghiên cứu khác đã kiểm tra các đặc điểm hành vi trong khi cố gắng giải thích cơ chế bệnh sinh cơ bản của chúng. Một phát hiện phổ biến là các mô hình chuột của SLOS cho thấy sự phát triển bất thường serotonergic, có thể chịu trách nhiệm một phần cho các hành vi tự kỷ được thấy trong SLOS. Mô hình chuột cũng đã được sử dụng để phát triển các kỹ thuật chẩn đoán; nhiều nghiên cứu đã kiểm tra chất đánh dấu sinh học là kết quả của oxy hóa của 7DHC, như DHCEO. Có khả năng là khi mô hình động vật được cải thiện, chúng sẽ dẫn đến nhiều khám phá hơn trong nghiên cứu SLOS.
Eponym
Hội chứng được đặt tên theo David Weyhe Smith (1926–1981), một bác sĩ nhi khoa Hoa Kỳ; Luc Lemli (1935–), một bác sĩ lâm sàng người Đức; và John Marius Opitz (1935–), một bác sĩ lâm sàng người Đức-Hoa Kỳ. Đây là những nhà nghiên cứu lần đầu tiên mô tả các triệu chứng của SLOS.
Tham khảo
Liên kết ngoài
Bản mẫu:Phakomatoses và dị tật bẩm sinh khác không được phân loại ở nơi khác Bản mẫu:Khiếm khuyết chuyển hóa steroid Bản mẫu:GHR