
Chuyển gen ngang là sự di chuyển của vật chất di truyền (DNA hoặc RNA) giữa các tế bào hoặc cơ thể sống cùng thế hệ, còn được gọi là truyền gen theo hàng ngang, để phân biệt với "chuyển gen theo hàng dọc" là quá trình kế thừa gen từ bố mẹ sang con cái thông qua sinh sản.
Thuật ngữ này dịch từ tiếng Anh là horizontal gene transfer (chuyển gen theo chiều ngang, hoặc chuyển gen hàng bên) viết tắt là HGT. Chuyển gen ngang phổ biến trong các loài vi khuẩn, còn trong tiến hoá thì quá trình này đóng vai trò rất quan trọng giúp các bộ gen của các loài rất khác nhau có thể trao đổi vật chất di truyền cho nhau mà không cần thông qua giao phối và thụ tinh.
Tổng quan
- HGT (chuyển gen ngang) đã được phát hiện từ lâu trong vi khuẩn học. Chẳng hạn: giữa hai cá thể trực khuẩn E. coli cùng thế hệ đã tiếp hợp, rồi qua lông giới tính mà DNA của cá thể này chuyển giao cho cá thể kia, tạo ra bộ gen mới không hề có giao phối và thụ tinh. Chính hiện tượng này là cơ chế phát tán tính chống thuốc kháng sinh ở rất nhiều loài vi khuẩn.
- Trong quá trình nội cộng sinh xa xưa, một loại vi khuẩn hiếu khí đã "chui" vào tế bào nhân thực cổ, từ đó hình thành nên ty thể ở hầu hết các tế bào sống ngày nay. Tương tự, sự chuyển gen ngang từ một loại vi khuẩn quang hợp xa xưa vào tổ tiên của thực vật đã hình thành nên lục lạp ở tất cả các cây xanh ngày nay (chi tiết vấn đề này xem ở trang nội cộng sinh). Những hiện tượng tiến hoá do nội cộng sinh này cũng là hiện tượng chuyển gen ngang.
- Tương tự thế, các gen của virut (DNA hoặc RNA) cũng có thể chuyển theo hàng ngang vào cơ thể sinh vật bậc cao, từ đó có thể góp phần tạo nên vốn gen của các sinh vật bậc cao này cho đến nay.
- Có thể do thói quen tư duy và cũng còn do sự phát triển của khoa học và công nghệ, nên hầu hết suy nghĩ về di truyền và kế thừa gen hiện nay đều đã chỉ tập trung vào "di truyền theo chiều dọc" qua các thế hệ của sinh vật, nhưng tầm quan trọng của việc chuyển gen ngang giữa trong quá trình tiến hoá không chỉ được thừa nhận mà việc nghiên cứu chúng phát triển mạnh.
Ý tưởng ban đầu của phương pháp chuyển gen nhân tạo (gene delivery) trong kỹ thuật di truyền có thể được gợi mở từ hiện tượng HGT của vi khuẩn trong tự nhiên.
Lược sử

- Năm 1928, Frederick Griffith đã công bố kết quả thí nghiệm, mà sau này thường được gọi bằng tên của ông, đó là thí nghiệm Griffith, được xem là bằng chứng đầu tiên về chuyển gen ngang. Gần 30 năm sau, một bài báo khoa học đã mô tả và chứng minh rằng việc "tiêm" virut vào vi khuẩn bạch hầu Corynebacterium diphtheriae đã tạo ra một chủng có độc lực từ một chủng không độc (năm 1951). Các nhà khoa học Nhật Bản đã mô tả lần đầu tiên về quá trình này trên một bài báo năm 1959 chứng minh sự chuyển gen chống kháng sinh giữa các loài vi khuẩn, từ đó gây ra sự "lây lan" vi khuẩn "nhờn thuốc". Khi bản chất của gen đã được hiểu rõ hơn, vào những năm 1980, thì Syvanen dự đoán rằng sự chuyển gen ngang tham gia vào việc quá trình tiến hóa của sinh giới kể từ khi sự sống bắt đầu trên Trái Đất. Sau đó, nhiều nhà khoa học cũng đồng ý với dự đoán này và có bằng chứng chứng tỏ chuyển gen ngang là một cơ chế tiến hóa quan trọng trong quá trình tiến hóa.
- Tiếp theo, sự chuyển gen ngang - từ một vài sự kiện riêng lẻ và đã gây lạ lẫm - lại trở thành khá phổ biến. Chẳng hạn như khi ghép một cành của cây này sang thân của cây khác có thể xảy ra quá trình chuyển lục lạp giữa hai cây, qua đó thì DNA lục lạp được trao đổi. Điều tương tự cũng xảy ra với DNA ty thể và những hiện tượng đó có nghĩa là tế bào "lai" chứa bộ gen có thể phát sinh dạng mới, thậm chí một loài mới. Vết cắn từ côn trùng Reduviidae có thể, thông qua ký sinh trùng, lây nhiễm cho người mắc trypanosomal từ đó gây ra bệnh Chagas, và DNA của nó vào chèn được bộ gen của người. Có ý kiến cho rằng việc chuyển gen ngang: người - vi khuẩn - người đã đóng vai trò phát sinh gen gây ung thư.
- Mặc dù chưa có tổng kết chính thức đầy đủ về vai trò của HGT trong tiến hoá, nhưng số lượng bằng chứng ngày càng gia tăng, sự phổ biến và tầm quan trọng của HGT trong sự tiến hóa ngày càng nhiều nên nhà sinh học phân tử Peter Gogarten đã gọi HGT là "một dạng thức mới cho sinh học" (A New Paradigm for Biology).
Cơ chế

Cơ chế HGT rất phức tạp, diễn ra ở mỗi nhóm loài không như nhau. Sau đây là một số.
- Biến nạp (transformation) là sự thay đổi vật chất di truyền của tế bào do tế bào "hấp thu" vật chất di truyền ngoại lai. Qua cơ chế này, vật liệu di truyền (là DNA hoặc RNA) từ loài này "xâm nhập" rồi "định cư" tại vật chủ nhận gen. Xem chi tiết hơn ở trang "Biến nạp" và "Kỹ thuật di truyền".
- Tải nạp (transduction) là quá trình DNA của virut được chuyển vào vi khuẩn, hoặc từ vi khuẩn này sang vi khuẩn khác nhờ virut (gồm cả thể thực khuẩn). Xem chi tiết hơn ở trang "Tải nạp" và "Kỹ thuật di truyền".
- Tiếp hợp vi khuẩn (bacterial conjugation) hoặc còn gọi là giao nạp (conjugation). Đây là quá trình chuyển DNA-plasmit từ tế bào vi khuẩn này (tế bào cho) sang tê bào vi khuẩn khác (tế bào nhận) qua quá trình gọi là tiếp hợp ở vi khuẩn (hình 2) được coi như là một kiểu sinh sản "hữu tính" nhờ lông tiếp hợp.
- Nhân tố chuyển gen (gene transfer agents) là các nhân tố giống như virut được tìm thấy ở nhóm vi khuẩn Rhodobacterales thuộc Alphaproteobacteria.
HGT ở vi sinh vật
Virut
Ở nhóm virut, có một loại tên là Mimivirus lây nhiễm amip. Một loại vi rút khác, được gọi là Sputnik ("virut vệ tinh") cũng lây nhiễm amip, nhưng nó không thể sinh sôi nảy nở trừ khi Mimivirus đã nhiễm vào cùng một tế bào chủ. Bộ gen của "virut vệ tinh" cho thấy đặc điểm sinh học của nó: trong số 13 gen của nó ít giống với bất kỳ gen nào khác đã biết, có 3 gen có liên quan mật thiết với các gen ở mimivirus, như là cần "cộng sinh" với chúng trong lịch sử tiến hóa, nên có tác giả gọi nó là virophague (virut kí sinh virut, như thể thực khuẩn). Điều này cho thấy rằng "virut vệ tinh" đã thực hiện chuyển gen theo chiều ngang giữa các virut, tương tự như phương thức các vi khuẩn chuyển gen ngang cho nhau.
Vi khuẩn
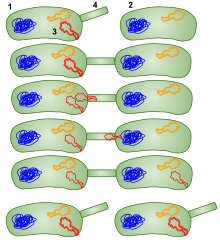
Ở vi khuẩn, biến nạp tự nhiên là sự thích nghi của vi khuẩn để chuyển DNA theo kiểu HGT. Nói chung, biến nạp có cơ chế phức tạp, đòi hỏi nhiều năng lượng. Để vi khuẩn có thể liên kết, tiếp nhận và tái tổ hợp DNA "ngoại lai" vào nhiễm sắc thể của nó, nó phải ở trạng thái sinh lý đặc biệt. Chẳng hạn, ở Bacillus subtilis trạng thái này đòi hỏi sự biểu hiện của khoảng 40 gen.
Xem thêm
- Sinh vật biến đổi gen.
- Nhân tố chuyển vị ngược.
- Nhân tố chuyển vị LTR.
- Thuyết nội cộng sinh.
- Cây phát sinh chủng loại.
- Cây phát sinh sự sống.
Tham khảo
- Citizendium:Horizontal gene transfer
- Citizendium:Horizontal gene transfer in prokaryotes
- Citizendium:Horizontal gene transfer in plants
- Citizendium:Horizontal gene transfer (History)
- Gyles, C; Boerlin, P (tháng 3 năm 2014). “Horizontally transferred genetic elements and their role in pathogenesis of bacterial disease”. Vet Pathol. 51 (2): 328–40. doi:10.1177/0300985813511131. PMID 24318976.
- – Papers by Dr Michael Syvanen on Horizontal Gene Transfer
-
Salzberg SL, White O, Peterson J, Eisen JA (tháng 6 năm 2001). “Microbial genes in the human genome: lateral transfer or gene loss?” (PDF). Science. 292 (5523): 1903–6. Bibcode:2001Sci...292.1903S. doi:10.1126/science.1061036. PMID 11358996. Bản gốc (PDF) lưu trữ ngày 1 tháng 9 năm 2006. Truy cập ngày 16 tháng 10 năm 2019.
About 40 genes were found to be exclusively shared by humans and bacteria and are candidate examples of horizontal transfer from bacteria to vertebrates. Gene loss combined with sample size effects and evolutionary rate variation provide an alternative, more biologically plausible explanation
- Qi, Z; Cui, Y; Fang, W; Ling, L; Chen, R (tháng 1 năm 2004). “Autosomal similarity revealed by eukaryotic genomic comparison”. Journal of Biological Physics. 30 (4): 305–12. doi:10.1007/s10867-004-0996-0. PMC 3456315. PMID 23345874.
- Woese CR (tháng 6 năm 2002). “On the evolution of cells”. Proc. Natl. Acad. Sci. U.S.A. 99 (13): 8742–7. Bibcode:2002PNAS...99.8742W. doi:10.1073/pnas.132266999. PMC 124369. PMID 12077305. This article seeks to shift the emphasis in early phylogenic adaptation from vertical to horizontal gene transfer. He uses the term "Darwinian Threshold" for the time of major transition of evolutionary mechanisms from mostly horizontal to mostly vertical transfer, and the "origin of speciation".
- Snel B, Bork P, Huynen MA (tháng 1 năm 1999). “Genome phylogeny based on gene content”. Nat. Genet. 21 (1): 108–10. doi:10.1038/5052. PMID 9916801. This article proposes using the presence or absence of a set of genes to infer phylogenies, in order to avoid confounding factors such as horizontal gene transfer.
- Webfocus in Nature with free review articles [1]
- Patil PB, Sonti RV (tháng 10 năm 2004). “Variation suggestive of horizontal gene transfer at a lipopolysaccharide (lps) biosynthetic locus in Xanthomonas oryzae pv. oryzae, the bacterial leaf blight pathogen of rice”. BMC Microbiol. 4 (1): 40. doi:10.1186/1471-2180-4-40. PMC 524487. PMID 15473911.
- Jin G, Nakhleh L, Snir S, Tuller T (tháng 11 năm 2006). “Maximum likelihood of phylogenetic networks”. Bioinformatics. 22 (21): 2604–11. doi:10.1093/bioinformatics/btl452. PMID 16928736. for a technique to decrease the impact of HGT events on maximum likelihood cladistical analyses.
- Horizontal Gene Transfer – A New Paradigm for Biology Lưu trữ 2012-07-21 tại Wayback Machine
- Horizontal Gene Transfer (page 334 of Molecular Genetics by Ulrich Melcher)
- Horizontal Gene Transfer at sciences.sdsu.edu
- Jain R, Rivera MC, Lake JA (tháng 3 năm 1999). “Horizontal gene transfer among genomes: The complexity hypothesis”. Proc. Natl. Acad. Sci. U.S.A. 96 (7): 3801–6. Bibcode:1999PNAS...96.3801J. doi:10.1073/pnas.96.7.3801. PMC 22375. PMID 10097118.
- PDF article on Horizontal Gene Transfer
- The New Yorker, ngày 12 tháng 7 năm 1999, pp. 44–61 "Smallpox knows how to make a mouse protein. How did smallpox learn that? 'The poxviruses are promiscuous at capturing genes from their hosts,' Esposito said. 'It tells you that smallpox was once inside a mouse or some other small rodent.'"
- Szpirer C, Top E, Couturier M, Mergeay M (ngày 1 tháng 12 năm 1999). “Retrotransfer or gene capture: a feature of conjugative plasmids, with ecological and evolutionary significance”. Microbiology. 145 (Pt 12): 3321–9. doi:10.1099/00221287-145-12-3321. PMID 10627031.
- GMO Safety: Results of research into horizontal gene transfer Lưu trữ 2011-07-21 tại Wayback Machine Can transgenes from genetically modified plants be absorbed by micro-organisms and spread in this way?
- Whitaker JW, McConkey GA, Westhead DR (2009). “The transferome of metabolic genes explored: analysis of the horizontal transfer of enzyme encoding genes in unicellular eukaryotes”. Genome Biology. 10 (4): R36. doi:10.1186/gb-2009-10-4-r36. PMC 2688927. PMID 19368726.