
| Hồi hải mã | |
|---|---|
 Người có hai hồi hải mã nằm ở hai bên bán cầu não. Hồi hải mã nằm ở thùy thái dương trong của não. Ở vị trí quan sát từ phía ngoài não, thùy trán ở bên trái, thùy chẩm ở bên phải và thùy thái dương, thùy đỉnh được cắt bỏ phần lớn để quan sát được hồi hải mã nằm ẩn ở phía dưới.
| |
 Hồi hải mã (màu hồng)
là một phần của hệ viền | |
| Chi tiết | |
| Một phần của | Thùy thái dương |
| Định danh | |
| Latinh | Hippocampus |
| MeSH | D006624 |
| NeuroName | 3157 |
| NeuroLex ID | birnlex_721 |
| TA | A14.1.09.321 |
| FMA | 275020 |
| Thuật ngữ giải phẫu thần kinh | |
Hồi hải mã (hay hải mã, cấu tạo hải mã, tiếng Anh: Hippocampus; tiếng Pháp: L'hippocampe, bắt nguồn từ tiếng Hy Lạp cổ đại ἱππόκαμπος, nghĩa là con cá ngựa) là thành phần quan trọng có trong não người và các động vật có xương sống khác. Con người và động vật có vú có hai hồi hải mã nằm ở hai bên não. Hồi hải mã là một phần của hệ viền (hệ limbic), có vai trò quan trọng trong việc củng cố trí nhớ (memory consolidation) từ trí nhớ ngắn hạn đến trí nhớ dài hạn và trí nhớ không gian cho phép điều hướng. Hồi hải mã nằm dưới vỏ đại não trong vỏ não nguyên thủy (allocortex), và ở bộ linh trưởng, hải mã nằm trong vùng trung gian thùy thái dương. Nó chứa hai phần lồng vào nhau: hải mã đích danh (hippocampus proprius, còn gọi là sừng Amon) và hồi răng (gyrus dentatus).
Khi mắc bệnh Alzheimer (và các dạng suy giảm trí nhớ khác), hải mã là một trong những vùng đầu tiên của não bị tổn thương, triệu chứng ban đầu là mất trí nhớ ngắn hạn và mất phương hướng. Tổn thương hải mã cũng có thể là do giảm oxy huyết, viêm não hoặc động kinh vùng trung gian thùy thái dương. Những người bị tổn thương hải mã rộng, ở hai bên có thể bị quên thuận chiều (anterograde amnesia): không có khả năng hình thành và lưu giữ những ký ức mới.
Do các tế bào thần kinh khác nhau được sắp xếp gọn gàng tầng tầng lớp lớp trong vùng hải mã, nên các nhà khoa học đã tạo nên mô hình khoa học để nghiên cứu sinh lý học thần kinh. Tính mềm dẻo synapse được biết đến trong quá trình điện thế hoá dài hạn (long-term potentiation, viết tắt: LTP) được phát hiện xảy ra ở hải mã và các nghiên cứu thường tập trung cấu trúc này. LTP được cho là một trong những cơ chế thần kinh chính mà theo đó các ký ức được lưu trữ trong não.
Lấy sinh vật mô hình là các loài gặm nhấm, có nhiều nghiên cứu chứng minh được rằng hải mã là một phần của hệ thống não chịu trách nhiệm cho trí nhớ không gian và điều hướng. Nhiều nơron trong hải mã của chuột phản ứng như tế bào chỗ (place cells, một phần của lớp tế bào tháp): các tế bào này tạo điện thế hoạt động khi động vật di chuyển qua một vị trí cụ thể thuộc môi trường xung quanh. Các tế bào chỗ ở hải mã tương tác mạnh mẽ với tế bào phụ trách phương hướng đầu con vật (head direction cells), có chức năng như một la bàn quán tính; và có lẽ chúng cũng tương tác với các tế bào lưới ở vỏ não nội khứu (entorhinal cortex) liền kề.
Lịch sử danh pháp

Bên trái: Hải mã ở người và vòm cung (hay thể tam giác não);
Bên phải: Con cá ngựa.
Nhà giải phẫu học người Ý tên là Julius Caesar Aranzi (1587) là người đầu tiên mô tả hải mã. Theo ông, cái "mào" này chạy dọc theo sàn của sừng thái dương của não thất bên, trông tựa như con tằm nhưng sau khi quan sát kĩ thì cho là giống con cá ngựa (Latin hippocampus, xuất phát từ tiếng Hy Lạp ἱππόκαμπος, trong đó ἵἵππς nghĩa là "con ngựa" + κάμπος nghĩa là "con quái vật biển"). Nhà giải phẫu học người Đức Duvernoy (1729) là người đầu tiên vẽ minh họa cấu trúc giải phẫu của hải mã, cũng phân vân không rõ nó giống "con cá ngựa" hay "con tằm". Năm 1732, nhà giải phẫu học người Đan Mạch Jacob Winsløw đề xuất danh pháp "sừng cừu đực" (Ram's horn). Một thập kỷ sau, bác sĩ phẫu thuật de Garengeot (đồng nghiệp tại Paris của Winsløw), đã sử dụng thuật ngữ "corne d'Ammon" - sừng Amon (lấy tên một vị thần Ai Cập cổ đại hay xuất hiện dưới hình dáng một người đàn ông với cái đầu cừu đực). Do vậy ở danh pháp giải phẫu tiếng Pháp, người ta viết tắt hải mã là CA.
Thuật ngữ chân hải mã (đầu sừng Amon) xuất hiện năm 1672, khi Diemerbroeck so sánh với hình dạng của cẳng chân hải mã trong truyền thuyết gập lại, trên chân có màng. Đây là một thủy quái mình ngựa đuôi cá. Giai đoạn sau này hải mã được mô tả là chân hải mã lớn (pes hippocampi major), còn chân hải mã bé (pes hippocampi minor) là chỗ phình ở ngay liền kề, nằm ở sừng chẩm, sau đó đặt lại tên là cựa chim (calcar avis). Tuy vậy, năm 1786, nhà giải phẫu học người Pháp Félix Vicq-d'Azyr đã hệ thống hóa, đặt lại tên hải mã là hải mã lớn, còn cựa chim là hải mã nhỏ. Năm 1779, Mayer viết nhầm thuật ngữ giải phẫu là hippopotamus (con hà mã) và một số tác giả khác tiếp tục sử dụng thuật ngữ nhầm này cho đến khi Karl Friedrich Burdach giải quyết lỗi danh pháp vào năm 1829. Năm 1861, Thomas Henry Huxley và Richard Owen tranh cãi gay gắt về sự tiến hóa của loài người dựa trên bằng chứng là cấu trúc "hải mã bé", được người đời gọi là Tranh cãi hải mã (Hippocampus-Debatte). Tuy vậy, trong sách giáo khoa giải phẫu không sử dụng thuật ngữ hải mã bé, và thuật ngữ này chính thức không được công nhận trong hệ thống danh pháp giải phẫu quốc tế Nomina Anatomica xuất bản năm 1895. Ngày nay, cấu trúc giải phẫu này chỉ gọi đơn thuần là hải mã (hay cấu trúc hải mã), còn thuật ngữ sừng Amon (Cornu Ammonis, tiếng Pháp: corne d'Ammon ) tồn tại trong tên viết tắt các diện hải mã thứ cấp (CA1 - CA4).
Liên quan đến hệ viền
Hệ viền (hay hệ limbic) do Paul MacLean phát hiện vào năm 1952 trong khi mô tả các cấu trúc ở chỗ viền của vỏ não (theo tiếng Latin, Limbus có nghĩa là bờ), gồm: hải mã, vỏ não đai, vỏ não khứu giác và hạch hạnh nhân. Paul MacLean sau đó cho rằng các cấu trúc viền là thần kinh nền tảng chi phối cảm xúc. Hồi hải mã có liên quan về mặt giải phẫu với các bộ phận có liên quan đến hành vi cảm xúc như nhân vách (septal nuclei), thể vú vùng dưới đồi và phức hợp nhân trước đồi thị (anterior nuclei of thalamus). Các cấu trúc này thường được chấp nhận là một phần của hệ viền.
Giải phẫu học


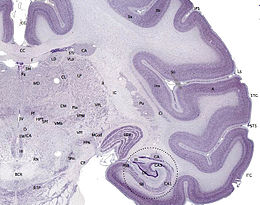
Hải mã được coi như là một dải mô chất xám, nằm trên nền sừng thái dương của não thất bên. Dải này cũng có thể được xem như là một nếp lộn vào trong của cổ vỏ não vào vùng trung gian thùy thái dương. Hồi hải mã chỉ có thể quan sát được trong phẫu tích vì nó bị hồi cạnh hải mã che lấp. Vỏ não chỗ này từ sáu lớp ban đầu giảm còn ba hoặc bốn lớp tạo nên vùng hải mã.
Thuật ngữ thành tạo hải mã (hippocampal formation) được sử dụng để chỉ hải mã đích danh (hippocampus proper) và các bộ phận liên quan của nó. Tuy nhiên, chưa thống nhất những cấu trúc giải phẫu bao gồm trong hai thuật ngữ này. Đôi khi, hải mã bao gồm hồi răng (dentate gyrus) và giá hải mã (subiculum). Một số tài liệu tham khảo cho rằng thành tạo hải mã bao gồm hồi răng và giá hải mã, một số tài liệu khác cho rằng thành tạo hải mã còn gồm cả tiền giá (presubiculum), cận giá (parasubiculum) và vỏ não nội khứu (entorhinal cortex). Ở tất cả các động vật có vú, đường đi thần kinh ở thành tạo hải mã là khá giống nhau.
Hồi hải mã bao gồm hồi răng có hình dạng giống chiếc ống uốn cong, được các nhà giải phẫu học so sánh như hình ảnh con cá ngựa và hình ảnh sừng cừu đực (Cornu Ammonis). Sử dụng chữ viết tắt CA để đọc danh pháp diện hải mã CA1, CA2, CA3 và CA4. Hải mã là một khu vực nơi vỏ não thu hẹp, tạo thành một lớp tế bào chóp dày đặc, cuộn tròn một cách chặt chẽ tạo thành hình chữ U. Diện CA4 là một viền của chữ "U" nêu trên, khu trú ở chỗ nối tiếp giữa hải mã với hồi răng, song ở trong rốn (hilus) của hồi răng. Hồi hải mã có phần trước và phần sau (ở linh trưởng) hoặc phần bụng và phần lưng ở các động vật khác. Cả hai phần có thành phần giống nhau nhưng thuộc các mạch nơron khác nhau. Ở chuột, hai hải mã trông giống như hai quả chuối nối với nhau ở mép sừng Amon. Ở linh trưởng, phần dưới của hải mã, gần nền thùy thái dương, rộng hơn nhiều so với phần trên. Tức là trong mặt phẳng cắt ngang, hải mã có thể có hình dạng khác nhau, tùy thuộc vào góc độ và vị trí của vết cắt.
Trong một thiết đồ cắt ngang hải mã bao gồm hồi răng, các lớp tế bào sẽ quan sát được. Hồi răng có ba lớp tế bào (hoặc bốn, nếu bao gồm các tế bào ở rốn - hilus). Các lớp từ bên ngoài vào trong: lớp phân tử, lớp phân tử trong, lớp hạt và rốn. Diện CA3 là một vùng tế bào lớn còn diện CA2 là một diện hỗn hợp, nằm trong hải mã. Diện CA1 là một vùng tế bào nhỏ, có thể tách thành hai lớp tế bào ở người, khu trú ở liền bề mặt giá hải mã-hải mã.
Luồng vào chính của cấu tạo hải mã là từ các tế bào của vỏ não nội khứu qua một đường chiếu phân tán gọi là đường xuyên (perforant path). Vỏ não nội khứu (entorhinal cortex, EC) là chỗ kết nối mạnh mẽ và đối ứng với nhiều cấu trúc vỏ não, dưới vỏ và thân não. Các nhân đồi thị khác khau, (từ đường giữa ngoài), nhân vách trong, các nhân trên núm vú (supramamillary nucleus) của hải mã, và cột giữa (raphe nuclei) nhân lục (locus coeruleus) của thân não đều gửi các sợi trục đến vỏ não nội khứu, do vậy nó đóng vai trò chuyển tiếp giữa vùng tân vỏ não (neocortex), các con đường kết nối và cấu tạo hải mã.
Vỏ não nội khứu nằm trong hồi cận hải mã. Hồi này che lấp hải mã, gồm vỏ não quanh khứu đóng vai trò quan trọng trong việc nhận dạng hình ảnh của các đối tượng phức tạp. Cũng có bằng chứng rằng hồi cận hải mã chi phối trí nhớ với những tác dụng có thể phân biệt với hải mã. Thật vậy, mất trí nhớ hoàn toàn chỉ xảy ra khi cả hải mã và cận hải mã bị tổn thương.
Sợi tới, sợi đi

Viết tắt:
DG: dentate gyrus (hồi răng);
Sub: subiculum (giá hải mã);
EC: entorhinal cortex (vỏ não nội khứu).
Luồng vào chính của cấu tạo hải mã đi qua vỏ não nội khứu (EC) trong khi luồng ra chính đi qua diện CA1 chiếu tới giá hải mã. Thông tin đến CA1 thông qua hai con đường chính, trực tiếp và gián tiếp. Các sợi trục từ vỏ não nội khứu bắt nguồn từ lớp III là nguồn gốc của đường xuyên (perforant path), hình thành các synapse trên các đuôi gai rất xa của các tế bào thần kinh CA1. Ngược lại, các sợi trục có nguồn gốc từ lớp II là nguồn gốc của con đường gián tiếp và thông tin đến CA1 thông qua mạch ba synapse. Trong phần đầu của con đường này, các sợi trục chiếu qua đường xuyên đến các tế bào hạt của hồi răng (synapse đầu tiên). Từ đó, thông tin theo sợi rêu đến CA3 (synapse thứ hai). Tiếp theo, các sợi trục CA3 (được gọi là bó Schaffer) rời khỏi phần sâu của soma, chiếu các sợi đến CA1 (synapse thứ ba). Các sợi trục từ CA1 sau đó chiếu trở lại vỏ não nội khứu, hoàn thành mạch thần kinh.
Tế bào rổ trong diện CA3 nhận sợi tới kích thích từ tế bào tháp và sau đó đưa ra phản hồi (điều hòa ngược) ức chế cho tế bào tháp. Sự ức chế hồi quy này là một vòng phản hồi đơn giản, giúp làm giảm các phản ứng kích thích ở hải mã. Tế bào chóp tạo kích thích hồi quy là một cơ chế quan trọng được tìm thấy trong một số vi mạch thần kinh xử lý bộ nhớ.
Một số kết nối thần kinh khác đóng vai trò quan trọng trong chức năng hải mã. Ngoài các sợi đi đến vỏ não nội khứu, hải mã còn có các sợi đi đến các vùng vỏ não khác bao gồm cả vỏ não trước trán. Một đầu ra chính đi qua vòm cung (fornix) đến vùng vách ngoài (lateral septal area) và đến thể núm vú của vùng dưới đồi (chỗ mà vòm cung liên kết với hải mã). Hồi hải mã nhận được sợi vào từ các hệ thống serotonin, norepinephrine, dopamine, và từ nhân Hensen (nucleus reuniens) của đồi thị đến diện CA1. Đường chiếu rất quan trọng đến từ nhân vách trong, gửi các sợi kích thích cholinergic (hay đường chiếu vách-hải mã tiết cholin - cholinergic septohippocampal projections) và gamma amino butyric acid (GABA) (sợi GABAergic) đến tất cả phần của hải mã. Các sợi vào từ nhân vách trong đóng vai trò chính trong việc kiểm soát trạng thái sinh lý của hải mã; phá hủy nhân này sẽ làm mất sóng theta hải mã và làm suy yếu nghiêm trọng một số loại trí nhớ.
Định khu

Các khu vực của hải mã khác biệt về chức năng và giải phẫu. Hải mã lưng (DH), hải mã bụng (VH) và hải mã trung gian chi phối các chức năng khác nhau, chiếu đến các con đường (pathway) khác nhau và có phân bố tế bào chỗ khác nhau. Hải mã lưng chi phối trí nhớ không gian, trí nhớ lời nói và ham muốn tìm hiểu thông tin. Sử dụng mô hình mê cung nước (Radial arm water maze), các tổn thương trong hải mã lưng được chứng minh là gây suy giảm trí nhớ không gian còn tổn thương hải mã bụng thì không bị. Sợi chiếu của nó đi đến nhân vách trong (medial septal nucleus) và nhân thể vú trên (supramammillary nucleus). Số lượng tế bào chỗ ở vùng hải mã lưng nhiều hơn cả vùng hải mã bụng và vùng hải mã trung gian cộng lại.
Vùng hải mã trung gian phân chia không rõ ràng với vùng hải mã bụng và lưng. Sử dụng các phương pháp theo dõi về sau (anterograde tracing), Cenquizca và Swanson (2007) đã đưa ra các đường chiếu cho hai vùng vỏ não khứu giác nguyên phát và vùng prelimbic (vùng tiền viền) của vỏ não trước trán trong. Đây là vùng có số lượng tế bào chỗ ít nhất. Hải mã bụng có chức năng trong điều kiện hóa sự sợ hãi và nhận thức tình cảm. Anagnostaras và đồng nghiệp (2002) đã chỉ ra rằng những thay đổi đối với hải mã bụng đã làm giảm lượng thông tin mà hải mã lưng và bụng gửi đến hạch nhân, do đó làm thay đổi tình trạng sợ hãi ở chuột. Trong lịch sử, giả thuyết được chấp nhận rộng rãi là hải mã có liên quan đến khứu giác. Giải thuyết này đã bị nghi ngờ, nhất là khi một loạt các nghiên cứu về giải phẫu không tìm thấy bất kỳ đường chiếu trực tiếp nào từ hành khứu giác đến hải mã. Tuy nhiên, công trình nghiên cứu sau này xác nhận rằng hành khứu giác có sợi chiếu vào phần bụng của vỏ não nội khứu ngoài, và diện CA1 trong vùng hải mã bụng gửi các sợi trục đến hành khứu giác chính,nhân khứu trước (anterior olfactory nucleus) và tới vỏ não khứu giác. Ngày nay rất ít chuyên gia tin rằng khứu giác là chức năng chính của hải mã, song vẫn có một số quan điểm về vai trò của hải mã trong trí nhớ mùi hương.
Chức năng
Các lý thuyết về chức năng hải mã
Trong những năm qua, ba chức năng chính của hải mã đã được nhắc đi nhắc lại trong các tài liệu, văn bản: kiểm soát ức chế (response inhibition), trí nhớ tình tiết (episodic memory) và nhận thức không gian. Lý thuyết ức chế hành vi (được John O'Keefe và Lynn Nadel mô tả) rất phổ biến ở những năm 1960. Lý thuyết này bắt nguồn phần lớn từ sự xác minh của hai quan sát thực nghiệm:
- Thứ nhất, động vật bị tổn thương vùng hải mã thị có xu hướng tăng động;
- Thứ hai, động vật bị tổn thương vùng hải mã thường gặp khó khăn trong việc học cách ức chế đáp ứng mà cá thể đã được dạy từ trước đó, đặc biệt là nếu đáp ứng theo yêu cầu giữ im lặng.
Jeffrey Gray phát triển quan sát này thành lý thuyết đầy đủ về vai trò của hải mã trong sự lo âu. Trong ba chức năng nêu trên, lý thuyết ức chế hiện giờ rất ít phổ biến.
Hải mã có chức năng quan trọng trong trí nhớ, và đã được thừa nhận trong lịch sử. Một báo cáo nổi tiếng của William Beecher Scoville và Brenda Milner mô tả kết quả cuộc phẫu thuật phá hủy hải mã cho bệnh nhân Henry Molaison ("Bệnh nhân H.M.", ông qua đời vào năm 2008) để làm giảm cơn động kinh. Kết quả khá bất ngờ: xuất hiện tình trạng quên thuận chiều và một phần quên ngược chiều; Molaison không còn khả năng hình thành những ký ức mới sau khi phẫu thuật và không thể nhớ bất kỳ sự kiện nào xảy ra ngay trước khi phẫu thuật, nhưng vẫn lưu lại những ký ức về những sự kiện xảy ra nhiều năm trước, nhất là từ thời thơ ấu. Trường hợp này thu hút các chuyên gia đến mức bệnh nhân Molaison trở thành chủ đề được nghiên cứu nhiều nhất trong lịch sử y học. Trong những năm sau đó, những bệnh nhân khác tổn thương hải mã có triệu chứng mất trí nhớ tương tự (có thể là do tai nạn hoặc bệnh tật) cũng đã được nghiên cứu, và hàng ngàn thí nghiệm nghiên cứu sinh lý học những thay đổi do hoạt động của các khớp nối synapse ở hải mã. Hiện nay có một quy ước phổ biến rằng hải mã có vai trò quan trọng trong bộ nhớ; tuy nhiên, bản chất chính xác của vai trò này vẫn còn gây tranh cãi.

Hải mã còn liên quan đến nhận thức không gian. Lý thuyết không gian ban đầu được O'Keefe và Nadel, người chịu ảnh hưởng bởi các lý thuyết của EC Tolman về "bản đồ nhận thức" ở người và động vật. O'Keefe và học trò Dostrovsky vào năm 1971 đã phát hiện ra các tế bào thần kinh ở hải mã chuột xuất hiện để thực hiện chức năng hoạt động có liên quan tới vị trí của chuột trong môi trường sống. Bất chấp sự hoài nghi của nhiều nhà nghiên cứu khác, O'Keefe và đồng nghiệp của ông, trong đó có Lynn Nadel, vẫn tiếp tục nghiên cứu vấn đề này. Hệ quả, cuốn sách Hải mã như một bản đồ nhận thức (The Hippocampus as a Cognitive Map) xuất bản năm 1978, có tầm ảnh hưởng sâu sắc. Hiện nay có quy ước rằng chức năng hồi hải mã đóng một vai trò quan trọng trong nhận thức không gian, nhưng cơ chế chi tiết như thế nào thì vẫn còn phải nghiên cứu.
Nghiên cứu sau này đã tập trung vào việc cố gắng thu hẹp tính rời rạc giữa hai quan điểm về chức năng hồi hải mã giữa trí nhớ và nhận thức không gian. Trong một số nghiên cứu, các chức năng này được tổng quát hóa đến mức gần hội tụ. Trong nỗ lực dung hòa hai quan điểm khác nhau, cần phải có tầm nhìn rộng hơn về chức năng hải mã. Theo khái niệm ban đầu của Tolman năm 1948, hải mã có khả năng tổ chức kinh nghiệm (bản đồ tâm thần), và định hướng hành vi. Đúng hơn, hải mã được coi là tham gia vào tất cả các khía cạnh của nhận thức. Vì vậy, chức năng của hải mã có thể được xem như là một hệ thống rộng, kết hợp cả trí nhớ và nhận thức không gian, đóng vai trò quan trọng trong một phạm vi rộng của bản đồ nhận thức. Tolman cho rằng điều này có liên quan đến việc chủ nghĩa hành vi có chủ đích sinh ra từ mục tiêu ban đầu, tức là ban đầu bạn phải nhận thức được mục đích và cơ chế phức tạp, sau đó chính những nhận thức này sẽ sinh ra hành vi.
Có đề xuất rằng hoạt động mạch xung (spiking activity) của các tế bào thần kinh vùng hải mã có liên quan mật thiết đến không gian, và có ý kiến cho rằng các cơ chế của trí nhớ và lập kế hoạch đều phát triển từ các cơ chế điều hướng; và rằng giải thuật nơron (neuronal algorithm) của hai cơ chế trên về cơ bản là giống nhau.
Nhiều nghiên cứu đã sử dụng các kỹ thuật chụp thần kinh như chụp cộng hưởng từ chức năng (fMRI) và vai trò chức năng trong xung đột chính-phụ đã được ghi nhận. Hải mã trước được xem là có liên quan đến việc ra quyết định theo phương pháp xử lý xung đột chính-phụ. Có ý kiến cho rằng trí nhớ, nhận thức không gian và các chức năng xử lý xung đột hoạt động cùng lúc và không loại trừ lẫn nhau.
Vai trò trí nhớ
Các nhà tâm lý học và khoa học thần kinh nói chung đều đồng ý rằng hải mã đóng vai trò quan trọng trong việc hình thành ký ức mới về những sự kiện kinh nghiệm (trí nhớ tình tiết hoặc trí nhớ tự truyện). Một phần của chức năng này có sự tham gia của hải mã trong việc phát hiện các sự kiện, địa điểm và kích thích mới. Một số nhà nghiên cứu coi hải mã là một phần của hệ thống trí nhớ của thùy thái dương trung gian chịu trách nhiệm chủ yếu cho trí nhớ quy nạp chung (ví dụ, những ký ức xác minh bằng lời nói, bao gồm trí nhớ ngữ nghĩa và trí nhớ tình tiết). Hồi hải mã cũng lưu trữ bối cảnh tình cảm từ hạch hạnh nhân. Đây là một phần của lý do tại sao mà khi ta trở lại địa điểm xảy ra một sự kiện có liên quan đến tình cảm, não bộ ta lại gợi lên cảm xúc đó. Có mối liên hệ tình cảm sâu sắc giữa trí nhớ và địa điểm.
Do tính đối xứng hai bên, não có hai hải mã nằm ở hai bán cầu đại não. Nếu tổn thương hải mã một bên, cấu trúc vẫn nguyên vẹn ở bên kia thì não vẫn duy trì chức năng bộ nhớ gần như bình thường. Tổn thương nghiêm trọng đối với hải mã ở cả hai bán cầu dẫn đến những khó khăn trong việc hình thành những ký ức mới (quên thuận chiều, anterograde amnesia) và cũng thường ảnh hưởng đến những ký ức hình thành trước khi tổn thương xảy ra (quên ngược chiều, retrograde amnesia). Mặc dù hiệu ứng quên về trước thường ảnh hưởng đến ký ức nhiều năm trước khi bị tổn thương não, song trong một số trường hợp những ký ức cũ vẫn còn. Việc lưu giữ những ký ức cũ này dẫn đến ý tưởng rằng có sự củng cố trí nhớ (memory consolidation) theo thời gian liên quan đến việc "di chuyển" những ký ức từ hải mã đến các phần khác của não. Nhiều thí nghiệm cấy ghép tế bào nội mô của tế bào hải mã ở loài linh trưởng có tổn thương thần kinh hải mã cho thấy rằng hải mã là cấu trúc cần thiết cho sự hình thành và hồi tưởng lại ký ức, nhưng không mang chức năng lưu trữ ký ức.
Tổn thương hải mã không ảnh hưởng đến một số loại trí nhớ, chẳng hạn như khả năng học các kỹ năng mới (chơi một nhạc cụ hoặc giải câu đố chẳng hạn). Thực tế này cho thấy những khả năng như vậy phụ thuộc vào các loại trí nhớ khác nhau (trí nhớ phương thức - procedural memory) và các định khu của não. Hơn nữa, bệnh nhân mất trí nhớ thường thể hiện trí nhớ "ngầm" cho các trải nghiệm cuộc sống ngay cả khi không còn kiến thức có chủ ý. Ví dụ, yêu cầu bệnh nhân tìm khuôn mặt nào trong hai khuôn mặt nhìn thấy gần đây nhất thì họ vẫn trả lời phần lớn là chính xác mặc dù họ khẳng định là chưa bao giờ thấy khuôn mặt. Một số nhà nghiên cứu phân biệt giữa hồi ức có ý thức liên quan đến hải mã và sự quen thuộc liên quan các phần của vùng thái dương trung gian.
Khi chuột phải trải qua quá trình học tập căng thẳng, chúng có thể giữ lại trí nhớ suốt đời về sự kiện này ngay cả sau một quá trình huấn luyện. Trí nhớ của một sự kiện có lẽ được lưu trữ đầu tiên ở hải mã, nhưng sự lưu trữ này là tạm thời. Phần lớn việc lưu trữ lâu dài của trí nhớ diễn ra ở vỏ não đai trước. Khi một quá trình học tập căng thẳng như vậy được áp dụng và thực nghiệm, hơn 5.000 vùng DNA methyl hóa xuất hiện trong bộ gen nơron ở hải mã của chuột trong 1 giờ và 24 giờ sau khi rèn luyện. Những thay đổi trong mô hình methyl hóa xảy ra ở nhiều gen được điều hoà xuống (down-regulated), do hình thành các vị trí 5-methylcytosine mới trong vùng giàu CpG của bộ gen. Hơn nữa, nhiều gen khác đã được điều hoà lên (up-regulated) do loại bỏ các nhóm methyl khỏi 5-methylcytosine (5mC) hiện có trong DNA. Một số protein hoạt động sẽ demethyl hóa 5mC, gồm enzyme Tet methylcytosine dioxygenase 1, các enzyme của con đường cắt và sửa base DNA (base excision repair) (xem Di truyền biểu sinh trong học tập và trí nhớ).
Vai trò trong trí nhớ không gian và điều hướng

Các nghiên cứu trên chuột di chuyển tự do cho thấy nhiều nơron hải mã có chức năng như các tế bào chỗ (place cell) tập hợp trong trường tế bào chỗ (place field) và những đánh dấu sự phóng điện do điện thế hoạt động khi con vật đi qua một địa điểm cụ thể. Hoạt động thần kinh liên quan đến vị trí của hải mã cũng được chứng minh ở những con khỉ được di chuyển xung quanh một căn phòng so với những con khỉ bị trói trên ghế. Tuy nhiên, sự phóng điện của các tế bào chỗ liên quan đến địa điểm con khỉ đang tìm kiếm hơn là vị trí thực tế của nó trong phòng. Trong nhiều năm, nhiều nghiên cứu đã được thực hiện về đáp ứng vị trí ở loài gặm nhấm đã cung cấp một lượng lớn thông tin. Đáp ứng của tế bào chỗ được thể hiện bởi tế bào chóp trong hải mã và bởi tế bào hạt trong hồi răng. Một lượng nhỏ các tế bào gây ức chế nơron trung gian (interneuron) và có sự biến đổi hoạt động thần kinh liên quan đến vị trí thể hiện qua việc nhịp phóng điện (firing rate) yếu hơn nhiều. Như vậy, các tế bào nằm cạnh nhau trong hải mã có mô hình phóng điện theo không gian là không tương quan (uncorrelated). Tế bào chỗ gần như không hoạt động khi con chuột di chuyển bên ngoài trường tế bào chỗ nhưng có thể đạt tốc độ duy trì lên tới 40 Hz, khi chuột ở gần trung tâm mô hình. Hoạt động thần kinh được lấy mẫu từ việc chọn ngẫu nhiên 30 đến 40 tế bào chỗ giúp mang đủ thông tin, cho phép tái tạo vị trí của chuột với độ tin cậy cao. Kích thước của các trường tế bào chỗ thay đổi theo gradient dọc theo chiều dài của hải mã, với các tế bào ở đầu phần lưng hải mã có trường nhỏ nhất, các tế bào gần giữa có trường lớn hơn và các tế bào ở phần lưng có trường bao phủ toàn bộ. Trong một số trường hợp, tốc độ phóng điện của các tế bào vùng hải mã không chỉ phụ thuộc vào vị trí mà còn phụ thuộc vào hướng di chuyển của con chuột, đích đến mà nó đang di chuyển hoặc các biến cố xảy ra khác. Việc phóng điện của các tế bào chỗ liên quan đến sóng theta cục bộ, một quá trình có tên là giai đoạn tuế sai (hay giai đoạn tiến động - phase precession).
Ở người, trong một nghiên cứu về bệnh nhân bị động kinh kháng thuốc (động kinh kháng trị), các tế bào có mô hình phóng điện theo vị trí cụ thể. Bệnh nhân trải qua thủ tục phẫu thuật xâm lấn để khoanh vùng nguồn gây ra cơn động kinh, nhằm phẫu thuật cắt bỏ. Họ được cấy điện cực chẩn đoán vào hải mã, sau đó sử dụng máy tính để ghi lại sự di chuyển trong một thành phố thực tế ảo. Các nghiên cứu ảnh chụp não về sự hoạt động của hải mã có liên quan đến tác dụng điều hướng. Một nghiên cứu đã được thực hiện trên các tài xế lái xe taxi tại London. Để vượt qua bài kiểm tra nghiêm ngặt tên là The Knowledge (Kiến thức) và lấy giấy phép lái xe, các tài xế phải tìm hiểu và nhớ một số lượng lớn địa điểm và tuyến đường nhanh nhất khi di chuyển. Nghiên cứu trên đã cho thấy phần sau của hải mã ở tài xế có kích thước lớn hơn so với người bình thường, và có mối tương quan chặt chẽ giữa tuổi nghề với sự gia tăng kích thước phần này. Tuy nhiên tổng thể tích của hải mã không thay đổi, vì phần sau gia tăng kích thước thì phần trước sẽ phải bù vào, kích thước giảm tương đối. Không có báo cáo về tác dụng phụ xuất phát từ sự chênh lệch tỷ lệ hải mã. Một nghiên cứu khác cho thấy ở những người khiếm thị, phần trước hải mã lại lớn hơn và phần sau hải mã nhỏ hơn so với người bình thường.
Ở não có một số loại tế bào được gọi là tế bào điều hướng. Chúng hoặc là phân bố ở trong hải mã, hoặc phân bố ngoài vùng nhưng có sự kết nối mạnh mẽ với hải mã, chẳng hạn như các tế bào tốc độ (speed cells) có trong vỏ não nội khứu trong. Các tế bào này cùng nhau tạo thành một mạng lưới có vai trò trí nhớ không gian. Loại tế bào tốc độ đầu tiên được phát hiện vào những năm 1970 là các tế bào chỗ, tiền đề cho ý nghĩ về cách hoạt động của hải mã đáp ứng thần kinh với môi trường trong bản đồ nhận thức. Khi hải mã bị rối loạn chức năng, điều hướng bị ảnh hưởng; bệnh nhân gặp khó khăn trong việc ghi nhớ đường đi đến một địa điểm và không thể đi xa. Đi lạc là một triệu chứng phổ biến của chứng hay quên. Nghiên cứu với động vật chỉ ra rằng hải mã nguyên vẹn giúp tạo trí nhớ ban đầu và duy trì lâu dài một số hành động liên quan đến trí nhớ không gian, đặc biệt là những hành động đòi hỏi phải tìm đường đến một mục tiêu được giấu kín. Các tế bào khác được phát hiện trong não của loài gặm nhấm nằm trong hải mã hoặc vỏ não, đó là các tế bào phụ trách phương hướng đầu con vật (head direction cell), các tế bào lưới và tế bào diềm (boundary cells). Các tế bào tốc độ cung cấp các sợi vào cho tế bào lưới hải mã.
Vai trò trong xử lý xung đột chính-phụ
Xung đột chính-phụ (approach-avoidance conflict) xảy ra khi tình huống đưa ra có thể là phần thưởng hay trừng phạt, và việc đưa ra quyết định hành động có liên quan đến sự lo lắng. Chụp fMRI trong các nghiên cứu xung đột chính-phụ đã tìm ra bằng chứng cho rằng: không thể lấy vai trò trí nhớ dài hạn hoặc nhận thức không gian của hải mã để giải thích cho vai trò xử lý xung đột. Các phát hiện tổng thể cho thấy rằng phần trước hải mã rất nhạy cảm với xung đột. Đây có thể là một phần của mạng lưới vỏ não và dưới vỏ não lớn, được coi là một phần quan trọng trong việc đưa ra quyết định trong điều kiện không chắc chắn.
Một đánh giá làm cơ sở tham chiếu cho nghiên cứu về sự liên quan của hải mã trong các hành vi xung đột. Tác giả cho rằng việc tìm hiểu cách xử lý xung đột liên quan đến chức năng điều hướng và trí nhớ không gian và cách thức mà tất cả các chức năng này không được loại trừ lẫn nhau vẫn còn nhiều trở ngại khó khăn.
Điện não đồ

Hải mã cho thấy hai "chế độ" hoạt động chính, mỗi chế độ liên quan đến một mô hình hoạt động của quần thể nơron (neuronal ensemble) và sóng hoạt động điện riêng biệt được đo bằng điện não đồ (EEG). Các "chế độ" này được đặt tên theo các đường đi trên bản EEG liên kết với chúng: theta và hoạt động bất thường biên độ lớn (LIA). Các đặc điểm chính được mô tả dưới đây được nghiên cứu trên loài động vật được nghiên cứu rộng rãi nhất, chuột.
Chế độ sóng theta xuất hiện trong trạng thái hoạt động để cảnh báo hành vi (đặc biệt là vận động) và trong giấc ngủ mắt chuyển động nhanh REM. Ở chế độ sóng theta, điện não đồ bị chi phối bởi các sóng lớn thông thường với dải tần từ 6 đến 9 Hz và các nhóm nơron chính vùng hải mã (tế bào chóp và tế bào hạt) cho thấy hoạt động quần thể nơron khá thưa thớt, chứng tỏ rằng trong bất kỳ khoảng thời gian ngắn nào, phần lớn tế bào đều ở trạng thái im lặng, trong khi phần nhỏ tế bào còn lại phóng điện thế với tốc độ tương đối cao, có thể lên tới 50 gai trong một giây. Mỗi tế bào hoạt động thường duy trì hoạt động trong nửa giây đến vài giây. Khi nghiên cứu trên chuột, các tế bào hoạt động luân phiên nhau, tỷ lệ phần trăm tế bào hoạt động rất ít thay đổi. Trong nhiều tình huống, hoạt động của tế bào được xác định chủ yếu bởi vị trí không gian của động vật, nhưng các biến cố hành vi cũng có ảnh hưởng rõ ràng.
Chế độ hoạt động bất thường biên độ lớn (LIA) xuất hiện trong giấc ngủ sóng chậm (không mơ màng) và cả trong trạng thái thức giấc nhưng bất động như nghỉ ngơi hoặc ăn uống. Trong chế độ LIA, điện não đồ bị chi phối bởi các sóng nhọn kéo dài trong 25–50 miligiây (ms). Sóng nhọn tạo ra theo bộ, mỗi bộ chứa tối đa 5 hoặc nhiều sóng nhọn riêng lẻ và kéo dài tới 500 ms. Hình dạng gai của nơron hải mã có mối tương quan cao với hình dạng sóng nhọn. Hầu hết nơron giảm tốc độ phóng điện giữa các sóng nhọn; tuy nhiên, trong một đợt sóng nhọn, tốc độ phóng tăng lên đáng kể ở 10% quần thể nơron vùng hải mã.
Hai chế độ hoạt động vùng hải mã này quan sát ở linh trưởng và chuột, tuy nhiên sóng theta ở linh trưởng không rõ ràng. Sóng nhọn và những thay đổi phụ thuộc trạng thái trong hoạt động quần thể nơron vẫn quan sát rõ.
Nhịp theta

Sóng theta được tạo ra chủ yếu bởi các lớp thần kinh dày đặc của vỏ não nội khứu, diện CA3 và sợi nhánh của tế bào chóp. Sóng theta là một trong những tín hiệu lớn nhất được thấy trên điện não đồ và gọi là nhịp theta hải mã. Trong một số trường hợp, điện não đồ bị chi phối bởi các sóng thường ở dải tần 3 đến 10 Hz, thường tiếp tục trong nhiều giây. Chúng phản ánh điện thế màng dưới ngưỡng và điều chỉnh mạnh mẽ đỉnh gai ở các nơron hải mã và đồng bộ hóa trên hải mã theo mô hình sóng di chuyển.Mạch ba synapse (trisynaptic circuit) là sự chuyển tiếp của dẫn truyền thần kinh ở vùng hải mã tương tác với nhiều vùng não. Từ các nghiên cứu trên loài gặm nhấm, có đề xuất cho rằng mạch ba synapse tạo ra nhịp theta hải mã.
Nhịp theta thấy rất rõ ràng ở thỏ, loài gặm nhấm, mèo và chó. Liệu nhịp theta thấy được ở các loài linh trưởng vẫn còn chưa rõ ràng. Ở chuột (động vật được nghiên cứu rộng rãi nhất), nhìn thấy nhịp theta khi thỏa mãn hai điều kiện:
- khi động vật đang đi hoặc tương tác tích cực với môi trường xung quanh.
- trong giấc ngủ mắt chuyển động nhanh REM.
Nhiều lý thuyết đã được đề xuất song chức năng của nhịp theta vẫn chưa được giải thích một cách thuyết phục. Giả thuyết phổ biến nhất liên quan đến học tập và trí nhớ. Một ví dụ: tại thời điểm kích thích nơron, nhịp theta định hình tác động của sự kích thích đó lên synapse. Tức là nhịp theta có thể ảnh hưởng đến học tập và trí nhớ phụ thuộc vào tính mềm dẻo synapse (synaptic plasticity). Tính chất này đã chứng minh được rằng các tổn thương của nhân đuôi trong (medial septum, nút trung tâm của hệ thống theta) gây ra sự gián đoạn nghiêm trọng của trí nhớ. Nhân đuôi trong không chỉ điều khiển nhịp theta; đây cũng là nguyên ủy chính của các sợi chiếu cholinergic đến hải mã. Tuy nhiên tính mềm dẻo nêu trên lại không làm sáng tỏ được rằng các tổn thương vùng nhân đuôi phát huy tác dụng bằng cách làm mất nhịp theta.
Sóng nhọn
Trong khi ngủ hoặc nghỉ ngơi, khi động vật không đáp ứng với môi trường xung quanh, điện não đồ hồi hải mã cho thấy một mô hình sóng chậm bất thường, biên độ lớn hơn một chút so với sóng theta. Mô hình này đôi khi bị gián đoạn bởi các đợt lớn được gọi là sóng nhọn. Hiện tượng này có liên quan với các đợt xuất hiện đỉnh gai trong điện não đồ kéo dài 50 đến 100 mili giây trong các tế bào chóp thuộc diện CA3 và diện CA1. Nó cũng liên quan tới các dao động điện não đồ tần số cao, thời gian ngắn, được gọi là "gợn" (ripple), với tần số trong phạm vi 150 đến 200 Hz ở chuột, và tồn tại cùng nhau gọi là sóng "Sharp wave-ripples" (SWRs). Sóng nhọn hay xuất hiện nhất trong khi ngủ với tốc độ trung bình khoảng 1 nhịp/giây (ở chuột) nhưng trong một mô hình thời gian rất bất thường. Sóng nhọn ít thấy hơn trong trạng thái thức không hoạt động và có cường độ nhỏ hơn. Sóng nhọn cũng quan sát thấy ở người và khỉ. Trong khỉ, sóng nhọn rất mạnh nhưng không thường xuyên như chuột.
Một trong những khía cạnh thú vị nhất của sóng nhọn là chúng có liên quan tới bộ nhớ. Wilson và McNaughton (1994) và nhiều nghiên cứu sau đó đã báo cáo rằng khi các tế bào chỗ ở hải mã có các trường phóng điện không gian chồng chéo (và do đó thường phóng ra gần như đồng thời). Chúng có xu hướng hiển thị lại các hoạt động tương quan trong khi ngủ sau khi đáp ứng hành vi. Sự tăng cường tính tương quan này, thường được gọi là tái kích hoạt, đã được tìm thấy xảy ra chủ yếu trong các sóng nhọn. Trên thực tế, người ta đã đề xuất rằng sóng nhọn là sự tái kích hoạt các mô hình hoạt động thần kinh được ghi nhớ trong quá trình hoạt động, được thúc đẩy bằng cách tăng cường các kết nối synapse trong vùng hải mã. Ý tưởng này tạo thành một tiền đề chính của lý thuyết "trí nhớ hai giai đoạn", được Buzsáki và nhiều người ủng hộ, trong đó đề xuất rằng các ký ức được lưu trữ trong vùng hải mã trong khi đáp ứng hành vi và sau đó được chuyển đến vùng tân vỏ não (neocortex) trong khi ngủ. Trong lý thuyết Hebbian, các sóng nhọn được xem là sự kích thích lặp đi lặp lại liên tục của các tế bào synapse cho và synapse nhận nhằm thúc đẩy các thay đổi synapse chỗ đích đến của sợi ra hồi hải mã, chính là vỏ não. Việc chặn sóng nhọn và gợn trong giấc ngủ hoặc khi bất động có thể gây trở ngại cho các ký ức thể hiện ở mức độ hành vi. Dù sao, tế bào chỗ mới ở diện CA1 có thể xuất hiện lại ngay cả sau khi sóng nhọn và gợn bị chặn đối với nhiệm vụ không đòi hỏi tư duy không gian.
Điện thế hóa dài hạn
Thời Ramon y Cajal (1852 - 1934), các nhà tâm lý học đã suy đoán rằng não lưu trữ trí nhớ bằng cách thay đổi sức mạnh kết nối giữa các nơron hoạt động đồng thời. Ý tưởng này đã được Donald Hebb chính thức hóa vào năm 1949, nhưng không giải thích được. Vào năm 1973, Tim Bliss và Terje Lømo mô tả một hiện tượng ở hải mã thỏ có đáp ứng với các thông số kỹ thuật mà Hebb đề xuất: sự thay đổi trong đáp ứng synapse gây ra bởi kích thích mạnh, ngắn và sự thay đổi này kéo dài hàng giờ, nhiều ngày hoặc lâu hơn. Hiện tượng này gọi là điện thế hóa dài hạn (long-term potential, LTP). Với tư cách là một cơ chế tiền năng để giải thích về trí nhớ dài hạn, LTP được nghiên cứu chuyên sâu và nghiệm ra được rất nhiều điều. Tuy nhiên, sự phức tạp và đa dạng của các tầng tín hiệu nội bào, vốn có thể kích thích LTP, chính là rào cản để nghiên cứu đầy đủ hơn.
Hải mã là một chi tiết giải phẫu đặc biệt thuận lợi để nghiên cứu LTP vì lớp nơron dày đặc và dễ dàng xác định. Nhưng các type thay đổi synapse phụ thuộc hoạt động cũng quan sát thấy ở nhiều vùng não khác. Dạng LTP hay nghiên cứu nhất đã được phát hiện trong diện CA1 hải mã, xảy ra tại các synapse trên gai răng (dendritic spine) và sử dụng chất dẫn truyền thần kinh glutamate. Sự thay đổi synapse phụ thuộc vào một loại thụ thể glutamate đặc biệt, thụ thể N -methyl-D-aspartate (NMDA), một thụ thể bề mặt tế bào có đặc tính đặc biệt là cho phép calci xâm nhập vào gai synapse nhận khi khử cực. Các loại thuốc can thiệp vào thụ thể NMDA ngăn chặn LTP có tác dụng chính đối với một số loại trí nhớ, đặc biệt là trí nhớ không gian. Chuột biến đổi gen được chỉnh sửa vật liệu di truyền nhằm vô hiệu hóa cơ chế LTP có bộ nhớ suy giảm nghiêm trọng.
Các loại rối loạn
Lão hóa
Những rối loạn liên quan đến tuổi tác như bệnh Alzheimer và các dạng suy giảm trí nhớ khác (trong đó rối loạn hồi hải mã là một trong những dấu hiệu sớm nhất) có tác động nghiêm trọng đến nhiều loại nhận thức, trong đó có cả trí nhớ. Ngay cả quá trình lão hóa bình thường cũng liên quan đến sự xuống cấp dần dần của một số dạng trí nhớ, bao gồm trí nhớ thường ngày và trí nhớ công việc (hay còn gọi là trí nhớ ngắn hạn). Vì hồi hải mã được cho là đóng vai trò quan trọng ở những chức năng liên quan đến trí nhớ, nên nhiều người đã cho rằng: những rối loạn về trí nhớ do lão hóa có thể bắt nguồn từ sự thoái hóa của vùng não này. Một số nghiên cứu ban đầu chỉ ra rằng những người cao tuổi bị mất một lượng đáng kể tế bào thần kinh ở hồi hải mã. Tuy nhiên, những nghiên cứu sau này với các kỹ thuật chính xác hơn cho thấy sự khác biệt là không đáng kể. Tương tự, một số nghiên cứu sử dụng kỹ thuật MRI cho thấy sự rút ngắn của hồi hải mã ở người cao tuổi, nhưng các nghiên cứu khác lại không tìm thấy đặc điểm này. Tuy nhiên, vẫn có một mối liên hệ đáng tin cậy giữa kích thước của hải mã và hiệu suất bộ nhớ; do vậy, nếu có sự rút ngắn vùng não vì lý do tuổi tác, hiệu suất trí nhớ sẽ bị suy giảm. Cũng có báo cáo rằng các nhiệm vụ nhớ có xu hướng tạo ra ít kích thích hơn ở người già so với người trẻ. Hơn nữa, một thử nghiệm đối chứng ngẫu nhiên được công bố vào năm 2011 cho thấy tập thể dục nhịp điệu có thể làm tăng kích thước của hồi hải mã ở người trưởng thành từ 55 đến 80 tuổi và cũng làm cải thiện trí nhớ không gian.
Căng thẳng
Hồi hải mã biểu hiện một lượng lớn thụ thể glucocorticoid, điều này khiến nó dễ bị tổn thương do căng thẳng lâu dài hơn hầu hết các vùng não khác. Có bằng chứng chỉ ra rằng những người trải qua các căng thẳng, chấn thương nghiêm trọng kéo dài bị teo hồi hải mã nhiều hơn các phần khác của não. Những ảnh hưởng này cũng xuất hiện trong hậu chấn tâm lý, và chúng có thể cũng liên quan đến hiện tượng teo hồi hải mã được báo cáo ở các bệnh tâm thần phân liệt và trầm cảm nặng. Một nghiên cứu gần đây cũng làm sáng tỏ việc teo hồi hải mã do hậu quả của trầm cảm, nhưng điều này có thể được cải thiện với thuốc chống trầm cảm, ngay cả khi những loại thuốc này không hiệu quả trong việc làm giảm các triệu chứng khác.
Căng thẳng mãn tính làm cho nồng độ glucocorticoid tăng cao, đáng chú ý là cortisol, được coi là một nguyên nhân gây teo tế bào thần kinh ở hồi hải mã. Sự teo này làm giảm thể tích của vùng não và cũng xuất hiện ở những bệnh nhân mắc hội chứng Cushing. Với những bệnh nhân mang hội chứng này, nồng độ cortisol cao thường là hệ quả do sử dụng các loại thuốc để chữa các triệu chứng khác. Các tế bào thần kinh cũng có thể bị mất đi do quá trình tạo mới thần kinh bị thoái hóa. Một yếu tố khác gây giảm thể tích hồi hải mã là sự co rút của các sợi nhánh trên tế bào thần kinh. Những sợi nhánh sẽ giảm cả về chiều dài và số lượng để đáp ứng với nồng độ glucocorticoid cao. Sự co rút sợi nhánh này là có thể đảo ngược. Sau khi điều trị bằng thuốc để giảm cortisol trong hội chứng Cushing, thể tích hồi hải mã được quan sát là có thể phục hồi tới 10%. Sự thay đổi này được coi là do sự phục hồi của các sợi nhánh. Sự phục hồi sợi nhánh này cũng có thể xảy ra khi căng thẳng biến mất. Tuy nhiên, một số nghiên cứu trên chuột cũng cho thấy rằng căng thẳng xảy ra ngay sau khi sinh có thể làm ảnh hưởng đến chức năng hồi hải mã trong suốt cuộc đời theo nhiều cách khác nhau.
Phản ứng đặc trưng giới tính đối với căng thẳng cũng ở chuột cũng được chứng minh là có ảnh hưởng đến hải mã. Căng thẳng mãn tính ở chuột đực cho thấy sự co rút sợi nhánh và suy giảm tế bào ở diện CA3 nhưng hiện tượng này không quan sát thấy ở con cái. Nguyên nhân cho sự khác biệt này có thể nằm ở hormone buồng trứng bảo vệ thần kinh. Ở chuột, các tổn thương DNA ở vùng hải mã tăng lên trong điều kiện căng thẳng.
Động kinh


Hồi hải mã là một trong số ít vùng não có thể tạo ra các tế bào thần kinh mới. Quá trình tạo mới thần kinh này được giới hạn trong hồi răng. Việc tạo ra các tế bào thần kinh mới này có thể được tăng cường nhờ luyện tập hoặc bị suy giảm bởi các cơn động kinh.
Động kinh vùng thùy thái dương có thể ảnh hưởng đến sự phát triển bình thường của các tế bào thần kinh mới và có thể gây tổn thương mô. Xơ cứng vùng hải mã là loại phổ biến nhất trong các tổn thương mô như vậy. Dù vậy, ta vẫn chưa biết rõ là tổn thương hải mã là nguyên nhân hay kết quả của các cơn động kinh. Tuy nhiên, khi gây động kinh nhân tạo nên động vật trong môi trường thí nghiệm, tổn thương hồi hải mã là kết quả thường gặp. Đây có thể là hậu quả do nồng độ các thụ thể glutamate có thể kích thích hải mã. Hưng phấn quá mức có thể làm đầu độc và gây chết tế bào. Sự tổn thương hồi hải mã trong các cơn động kinh cũng có thể có liên quan đến việc các tế bào thần kinh mới tiếp tục được tạo ra ở vùng này trong suốt cuộc đời và những bất thường trong quá trình này.
Tâm thần phân liệt
Nguyên nhân của chứng tâm thần phân liệt đến nay vẫn chưa được hiểu rõ, nhưng nhiều bất thường về cấu trúc não đã được ghi nhận. Những thay đổi liên quan đến vỏ não được nghiên cứu rất kĩ lưỡng và những ảnh hưởng đối với hồi hải mã cũng đã được mô tả. Nhiều báo cáo đã tìm thấy sự giảm kích thước của hồi hải mã ở những người bị tâm thần phân liệt. Hồi hải mã bên trái dường như bị ảnh hưởng nhiều hơn bên phải. Các nhà khoa học đồng tình rằng những thay đổi này là hậu quả của sự phát triển bất thường. Dù vậy, ta vẫn chưa biết rõ rằng liệu sự thay đổi hồi hải mã có vai trò gì trong việc gây ra các triệu chứng loạn thần, đặc điểm quan trọng nhất của chứng tâm thần phân liệt hay không. Trên cơ sở từ những thí nghiệm sử dụng động vật, có ý kiến cho rằng: rối loạn chức năng vùng hải mã có thể làm ảnh hưởng đến việc giải phóng dopamine trong hạch nền, gián tiếp ảnh hưởng đến sự tích hợp thông tin ở thùy não trước trán. Cũng có ý kiến cho rằng: rối loạn chức năng ở hồi hải mã có thể giải thích cho hiện tượng rối loạn trí nhớ dài hạn thường được ghi nhận ở các bệnh nhân tâm thần phân liệt.
Các nghiên cứu sử dụng kỹ thuật MRI cho thấy: những người bị tâm thần phân liệt có khối lượng não nói chung và não thất nhỏ hơn người bình thường, tuy nhiên các nhà nghiên cứu không biết rằng liệu sự teo này là do bệnh tâm thần phân liệt hay từ thuốc sử dụng để trị bệnh. Vùng hồi hải mã và đồi thị cũng cho thấy sự giảm thể tích, nhưng thể tích của globus pallidus lại tăng lên. Mô hình vỏ não ở các bệnh nhân này cũng bị thay đổi, thể tích và độ dày của lớp vỏ não, đặc biệt là ở thùy trán và thái dương, cũng được ghi nhận là giảm. Những biến đổi của não trong quá trình xuất hiện và tiến triển của bệnh củng cố cho giả thuyết rằng hệ thần kinh đã phát triển không bình thường ở những trường hợp này.
Mất trí nhớ toàn phần thoáng qua
Mất trí nhớ toàn phần thoáng qua sẽ khiến cho nạn nhân mất gần như hoàn toàn trí nhớ ngắn hạn trong một khoảng thời gian ngắn, thường xảy ra đột ngột. Nguyên nhân của dạng mất trí nhớ này được cho là do thiếu máu, động kinh, đau nửa đầu migraine và rối loạn lưu lượng máu tĩnh mạch não, làm thiếu máu cục bộ tại các cấu trúc như hồi hải mã, vùng não có liên quan mật thiết đến trí nhớ.
Những nguyên nhân kể trên đều chưa có bằng chứng khoa học ủng hộ. Tuy nhiên, các nghiên cứu sử dụng kỹ thuật MRI khuếch tán trên những nạn nhân trong vòng 12 đến 24 giờ sau khi xuất hiện các triệu chứng có cho thấy những tổn thương giống như những chấm nhỏ ở vùng hải mã. Những phát hiện này đã gợi ý rằng, các tế bào thần kinh vùng CA1 có thể sẽ dễ bị tổn thương do những căng thẳng trao đổi chất.
Hậu chấn tâm lý
Một số nghiên cứu cho thấy mối liên quan giữ việc giảm thể tích của hồi hải mã và stress sau khi xảy ra sang chấn (hậu chấn tâm lý hay PTSD). Một nghiên cứu được thực hiện trên những người lính Mỹ từng tham chiến tại Việt Nam với hậu chấn tâm lý cho thấy, thể tích hồi hải mã của họ nhỏ hơn 20% nếu so sánh với các cựu chiến binh khác không bị stress sau sang chấn. Tuy nhiên, phát hiện này không được tìm thấy ở những nạn nhân bị hậu chấn tâm lý sau khi xảy ra thảm kịch rơi máy bay ở triển lãm hàng không năm 1988 (Ramstein, Đức). Một trường hợp khác là những anh em sinh đôi không tham chiến của các cựu chiến binh Mỹ trong chiến tranh Việt Nam mắc PTSD cũng có hồi hải mã nhỏ hơn so với nhóm đối tượng đối chứng, đặt ra câu hỏi về bản chất của mối liên quan giữa hai hiện tượng này. Một nghiên cứu năm 2016 đã ủng hộ cho giả thuyết rằng thể tích hồi hải mã nhỏ sẽ làm tăng nguy cơ mắc hậu chấn tâm lý (chứ không phải stress sau sang chấn sẽ gây tổn thương hồi hải mã) và các phương pháp chữa trị cũng hiệu quả với những người có hồi hải mã lớn hơn.
Những động vật khác

Động vật có vú
Ở lớp Thú (Động vật có vú), từ các loài thuộc bộ Đơn huyệt như tachyglossidae đến các loài thuộc bộ Linh trưởng như người, hải mã đều có cấu tạo gần như nhau. Tỷ lệ kích thước hải mã so với cơ thể tăng lên đáng kể theo chiều hướng tiến hóa: bộ Linh trưởng tăng gấp hai lần so với bộ Đơn huyệt. Tuy nhiên, tỷ lệ kích thước hải mã lại không tăng nhanh như tỷ lệ kích thước tân vỏ não so với cơ thể. Do đó so với loài linh trưởng, hải mã ở loài gặm nhấm vẫn chiếm một phần lớn hơn nhiều trên lớp vỏ não. Ở người trưởng thành, thể tích hải mã mỗi bên não chiếm khoảng 3,0 đến 3,5 cm³ so với thể tích của tân vỏ não là 320 đến 420 cm³.
Có mối quan hệ chung giữa kích thước hải mã và trí nhớ không gian. Khi so sánh giữa các loài, những loài có trí nhớ không gian lớn hơn thì thể tích hồi hải mã có xu hướng lớn hơn. Sự khác biệt về giới tính cũng ảnh hưởng đến thể tích hải mã: ở những loài mà con đực và con cái có sự khác biệt rõ rệt về khả năng ghi nhớ không gian thì thể tích hải mã cũng có sự khác biệt.
Động vật có xương sống khác
Các loài không thuộc lớp thú không có cấu trúc hải mã, nhưng chúng có cơ quan tương đồng với nó. Như các nội dung đã nêu ở trên, về bản chất, hải mã là một phần của vỏ não nguyên thủy (allocortex). Chỉ các động vật lớp Thú có vỏ não phát triển đầy đủ, nhưng áo não (pallium) (cấu trúc nguyên thủy của hải mã) lại có mặt trong tất cả các động vật có xương sống, ngay cả ở những loài nguyên thủy như cá mút đá hoặc cá mixini. Áo não thường được chia thành ba khu vực: trong, ngoài và sau. Áo não trong tạo thành tiền thân của đồi hải mã. Nó không giống với hải mã khi quan sát vì các lớp không cuộn lại thành hình chữ S hoặc được bao bọc bởi hồi răng, nhưng sự tương đồng biểu hiện nhờ ái lực hóa học và chức năng hoạt động mạnh mẽ. Hiện nay có bằng chứng cho thấy những cấu trúc giống như đồi hải mã này có liên quan đến nhận thức không gian ở chim, bò sát và cá.
Chim
Ở các loài chim, cơ quan tương đồng đầy đủ đến mức hầu hết các nhà giải phẫu học đều gọi vùng áo não trong là "hải mã chim" (avian hippocampus). Rất nhiều loài chim có kỹ năng nhận biết không gian mạnh mẽ, đặc biệt là những loài cất giữ thức ăn. Có bằng chứng cho thấy những con chim cất giữ thức ăn có cấu tạo hải mã lớn hơn các loại chim khác và nếu bị tổn thương hải mã sẽ ảnh hưởng bất lợi tới trí nhớ không gian.
Cá
Ở các loài cá thì phức tạp hơn. So với các loại động vật có xương sống khác, ở phân thứ lớp Cá xương thật (chiếm phần lớn trong số các loài hiện có), não trước bị biến dạng: Hầu hết các nhà giải phẫu học thần kinh cho rằng não trước của các loài cá thuộc phân thứ lớp này, về bản chất, tựa như một chiếc tất lộn từ trong ra ngoài. Do đó, các cấu trúc nằm ở bên trong (bên cạnh não thất) đối với hầu hết các loài động vật có xương sống, lại được tìm thấy ở bên ngoài ở phân thứ lớp Cá xương thật và ngược lại. Hệ quả là áo não trong (vùng "hải mã") của động vật có xương sống tương ứng với áo não ngoài của cá. Một số loại cá (đặc biệt là cá vàng) đã được chứng minh bằng thực nghiệm có khả năng trí nhớ không gian mạnh mẽ, thậm chí hình thành "bản đồ nhận thức" về khu vực chúng sinh sống. Có bằng chứng cho thấy tổn thương áo não ngoài ảnh hưởng xấu tới trí nhớ không gian của cá. Người ta vẫn chưa biết liệu áo não ngoài có vai trò tương tự ở các loài động vật có xương sống nguyên thủy hơn, chẳng hạn như cá mập và cá đuối, hay thậm chí là cá mút đá và cá mixini.
Côn trùng và động vật thân mềm
Một số loại côn trùng và động vật thân mềm như bạch tuộc, cũng có khả năng học tập và điều hướng không gian mạnh mẽ, nhưng có vẻ hoạt động khác với hệ không gian của động vật có vú, vì vậy không lý do hợp lý nào để nghi ngờ chúng có nguồn gốc tiến hóa chung; cũng không có đủ sự tương đồng về cấu trúc não để xuất hiện chi tiết giải phẫu "giống như hải mã" ở những loài này. Tuy nhiên có đề xuất cho rằng thể cuống (mushroom bodies) của côn trùng có thể có chức năng tương tự như hải mã.
Hình ảnh
-
Hải mã ở chim bồ câu
-

Hình thái hải mã ở bốn loài thuộc bộ gặm nhấm
-

Vùng hải mã chính của khỉ marmoset
-

Phân bố số lượng tế bào giữa các loài
-

Số lượng tế bào so với tỷ lệ trọng lượng cơ thể ở loài gặm nhấm
-

Mất nơron diện CA1 do thiếu máu cục bộ ở chuột






Ghi chú
Tham khảo
- Aboitiz F, Morales D, Montiel J (tháng 10 năm 2003). “The evolutionary origin of the mammalian isocortex: towards an integrated developmental and functional approach” [Nguồn gốc tiến hóa của tân vỏ não ở động vật có vú: tiếp cận theo sự phát triển và chức năng]. The Behavioral and Brain Sciences. 26 (5): 535–52. doi:10.1017/S0140525X03000128. PMID 15179935.
- Amaral D, Lavenex P (2006). “Ch 3. Hippocampal Neuroanatomy”. Trong Andersen P, Morris R, Amaral D, Bliss T, O'Keefe J (biên tập). The Hippocampus Book [Sách về hải mã]. Oxford University Press. ISBN 978-0-19-510027-3.
- Anagnostaras SG, Gale GD, Fanselow MS (2002). “The hippocampus and Pavlovian fear conditioning: reply to Bast et al” [Hải mã và sự điều hòa sợ hãi phản xạ Pavlov: câu trả lời của Bast và đồng nghiệp] (PDF). Hippocampus. 12 (4): 561–565. doi:10.1002/hipo.10071. PMID 12201641. Bản gốc (PDF) lưu trữ ngày 16 tháng 2 năm 2005.
- Best PJ, White AM (1999). “Placing hippocampal single-unit studies in a historical context” ['bối cảnh lịch sử nghiên cứu từng vùng hải mã]. Hippocampus. 9 (4): 346–51. doi:10.1002/(SICI)1098-1063(1999)9:4<346::AID-HIPO2>3.0.CO;2-3. PMID 10495017.
- Bliss TV, Lomo T (tháng 7 năm 1973). “Long-lasting potentiation of synaptic transmission in the dentate area of the anaesthetized rabbit following stimulation of the perforant path” [Điện thế hóa dài hạn của dẫn truyền synapse tại hồi răng của thỏ gây mê sau khi kích thích đường xuyên]. The Journal of Physiology. 232 (2): 331–56. doi:10.1113/jphysiol.1973.sp010273. PMC 1350458. PMID 4727084.
- Boyer P, Phillips JL, Rousseau FL, Ilivitsky S (tháng 4 năm 2007). “Hippocampal abnormalities and memory deficits: new evidence of a strong pathophysiological link in schizophrenia” [Hải mã bất thường và suy giảm trí nhớ: bằng chứng mới về sự liên quan giữa sinh lý bệnh trong tâm thần phân liệt]. Brain Research Reviews. 54 (1): 92–112. doi:10.1016/j.brainresrev.2006.12.008. PMID 17306884.
- Broglio C, Gómez A, Durán E, Ocaña FM, Jiménez-Moya F, Rodríguez F, Salas C (tháng 9 năm 2005). “Hallmarks of a common forebrain vertebrate plan: specialized pallial areas for spatial, temporal and emotional memory in actinopterygian fish”. Brain Research Bulletin. 66 (4–6): 397–99. doi:10.1016/j.brainresbull.2005.03.021. PMID 16144602.
- Burke SN, Barnes CA (tháng 1 năm 2006). “Neural plasticity in the ageing brain” [Sự mềm dẻo thần kinh trong lão hóa não]. Nature Reviews. Neuroscience. 7 (1): 30–40. doi:10.1038/nrn1809. PMID 16371948.
- Buzsáki G (tháng 11 năm 1986). “Hippocampal sharp waves: their origin and significance” [Sóng nhọn hải mã: nguồn gốc và ý nghĩa]. Brain Research. 398 (2): 242–52. doi:10.1016/0006-8993(86)91483-6. PMID 3026567.
- Buzsáki G (1989). “Two-stage model of memory trace formation: a role for "noisy" brain states” [Mô hình hai giai đoạn hình thành dấu vết trí nhớ]. Neuroscience. 31 (3): 551–70. doi:10.1016/0306-4522(89)90423-5. PMID 2687720.
- Buzsáki G, Chen LS, Gage FH (1990). “Chapter 19 Chapter Spatial organization of physiological activity in the hippocampal region: Relevance to memory formation”. Spatial organization of physiological activity in the hippocampal region: relevance to memory formation [Tổ chức không gian của hoạt động sinh lý ở hải mã: sự liên quan đến hình thành trí nhớ]. Progress in Brain Research. 83. tr. 257–68. doi:10.1016/S0079-6123(08)61255-8. ISBN 9780444811493. PMID 2203100.
- Buzsáki G (tháng 1 năm 2002). “Theta oscillations in the hippocampus” [Dao động theta ở hải mã] (PDF). Neuron. 33 (3): 325–40. doi:10.1016/S0896-6273(02)00586-X. PMID 11832222. Bản gốc (PDF) lưu trữ ngày 25 tháng 6 năm 2008.
- Buzsáki, G (2006). Rhythms of the Brain [Nhịp não]. Oxford University Press. ISBN 978-0-19-530106-9.
- Ramón y Cajal S (1894). “The Croonian Lecture: La Fine Structure des Centres Nerveux” [Bài giảng Croonia: Cấu trúc tinh tế của trung tâm thần kinh]. Proceedings of the Royal Society. 55 (331–335): 444–68. Bibcode:1894RSPS...55..444C. doi:10.1098/rspl.1894.0063.
- Guoxiang Xiong, Hannah Metheny, Brian N. Johnson, và Akiva S. Cohen (2017). “A Comparison of Different Slicing Planes in Preservation of Major Hippocampal Pathway Fibers in the Mouse” [So sánh các mặt phẳng cắt khác nhau trong việc bảo tồn sợi chính trong con đường dẫn truyền vùng hải mã ở chuột]. NCBI. Bibcode:1894RSPS...55..444C. doi:10.3389/fnana.2017.00107. PMC 5696601. PMID 29201002.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- Campbell S, Macqueen G (tháng 11 năm 2004). “The role of the hippocampus in the pathophysiology of major depression” [Vai trò của hải mã trong sinh lý bệnh của bệnh trầm cảm]. Journal of Psychiatry & Neuroscience. 29 (6): 417–26. PMC 524959. PMID 15644983.
- Cantero JL, Atienza M, Stickgold R, Kahana MJ, Madsen JR, Kocsis B (tháng 11 năm 2003). “Sleep-dependent theta oscillations in the human hippocampus and neocortex” [Dao động theta phụ thuộc vào giấc ngủ ở hải mã và vùng tân vỏ não của con người]. The Journal of Neuroscience. 23 (34): 10897–903. doi:10.1523/JNEUROSCI.23-34-10897.2003. PMC 6740994. PMID 14645485.
- Carey, B (ngày 4 tháng 12 năm 2008). “H. M., an Unforgettable Amnesiac, Dies at 82” [H. M., bệnh nhân mất trí nhớ, qua đời ở tuổi 82]. The New York Times. Truy cập ngày 27 tháng 4 năm 2009.
- Chiu YC, Algase D, Whall A, Liang J, Liu HC, Lin KN, Wang PN (2004). “Getting lost: directed attention and executive functions in early Alzheimer's disease patients” [Đi lạc: sự chú ý phương hướng và chức năng điều hành ở bệnh nhân mắc bệnh Alzheimer giai đoạn sớm]. Dementia and Geriatric Cognitive Disorders. 17 (3): 174–80. doi:10.1159/000076353. PMID 14739541.
- Chang BS, Lowenstein DH (tháng 9 năm 2003). “Epilepsy” [Động kinh]. The New England Journal of Medicine. 349 (13): 1257–66. doi:10.1056/NEJMra022308. PMID 14507951.
- Cho RY, Gilbert A, Lewis DA (2005). “Ch 22. The neurobiology of schizophrenia”. Trong Charney DS, Nestler EJ (biên tập). Neurobiology of Mental Illness [Sinh học thần kinh của bệnh tâm thần]. Oxford University Press US. ISBN 978-0-19-518980-3.
- Cenquizca LA, Swanson LW (tháng 11 năm 2007). “Spatial organization of direct hippocampal field CA1 axonal projections to the rest of the cerebral cortex” [Tổ chức không gian của các sợi chiếu hải mã diện CA1 đến phần còn lại của vỏ não]. Brain Research Reviews. 56 (1): 1–26. doi:10.1016/j.brainresrev.2007.05.002. PMC 2171036. PMID 17559940.
- Clark RE, Broadbent NJ, Squire LR (2005). “Hippocampus and remote spatial memory in rats” [Hải mã và bộ nhớ không gian ở chuột]. Hippocampus. 15 (2): 260–72. doi:10.1002/hipo.20056. PMC 2754168. PMID 15523608.
- Colombo M, Broadbent N (tháng 6 năm 2000). “Is the avian hippocampus a functional homologue of the mammalian hippocampus?” [Hải mã chim liệu có sự tương đồng về chức năng so với hải mã động vật có vú?]. Neuroscience and Biobehavioral Reviews. 24 (4): 465–84. doi:10.1016/S0149-7634(00)00016-6. PMID 10817844.
- Cooke SF, Bliss TV (tháng 7 năm 2006). “Plasticity in the human central nervous system” [Tính mềm dẻo của hệ thần kinh trung ương người]. Brain. 129 (Pt 7): 1659–73. doi:10.1093/brain/awl082. PMID 16672292.
- de Olmos J, Hardy H, Heimer L (tháng 9 năm 1978). “The afferent connections of the main and the accessory olfactory bulb formations in the rat: an experimental HRP-study” [Các đường đi hướng tâm và sự hình thành hành khứu giác ở chuột: nghiên cứu thử nghiệm với enzyme peroxidase cải ngựa]. The Journal of Comparative Neurology. 181 (2): 213–244. doi:10.1002/cne.901810202. PMID 690266.
- Diana RA, Yonelinas AP, Ranganath C (tháng 9 năm 2007). “Imaging recollection and familiarity in the medial temporal lobe: a three-component model” [Hình ảnh các cấu trúc phụ trách hồi ức và sự quen thuộc ở thùy thái dương: mô hình ba thành phần]. Trends in Cognitive Sciences. 11 (9): 379–86. doi:10.1016/j.tics.2007.08.001. PMID 17707683.
- Duvernoy, HM (2005). “Introduction”. The Human Hippocampus [Hải mã người] (ấn bản 3). Berlin: Springer-Verlag. tr. 1. ISBN 978-3-540-23191-2.
- Eichenbaum H, Otto TA, Wible CG, Piper JM (1991). “Ch 7. Building a model of the hippocampus in olfaction and memory”. Trong Davis JL, Eichenbaum H (biên tập). Olfaction [Khứu giác]. MIT Press. ISBN 978-0-262-04124-9.
- Eichenbaum H, Cohen NJ (1993). Memory, Amnesia, and the Hippocampal System [Trí nhớ, sự mất trí nhớ và hệ hải mã]. MIT Press.
- Eichenbaum H, Yonelinas AP, Ranganath C (2007). “The medial temporal lobe and recognition memory” [Thùy thái dương trung gian và trí nhớ nhận dạng]. Annual Review of Neuroscience. 30: 123–52. doi:10.1146/annurev.neuro.30.051606.094328. PMC 2064941. PMID 17417939.
- Ekstrom AD, Kahana MJ, Caplan JB, Fields TA, Isham EA, Newman EL, Fried I (tháng 9 năm 2003). “Cellular networks underlying human spatial navigation” [Mạng lưới tế bào chi phối điều hướng không gian của người] (PDF). Nature. 425 (6954): 184–88. Bibcode:2003Natur.425..184E. doi:10.1038/nature01964. PMID 12968182.
- Erickson KI, và đồng nghiệp (tháng 2 năm 2011). “Exercise training increases size of hippocampus and improves memory (Luyện tập làm tăng kích thước hải mã và cải thiện trí nhớ)”. Proceedings of the National Academy of Sciences of the United States of America. 108 (7): 3017–3022. Bibcode:2011PNAS..108.3017E. doi:10.1073/pnas.1015950108. PMC 3041121. PMID 21282661.
- Fanselow MS, Dong HW (tháng 1 năm 2010). “Are the dorsal and ventral hippocampus functionally distinct structures?” [Liệu hải mã lưng và hải mã bụng là hai cấu trúc có chức năng khác nhau?]. Neuron. 65 (1): 7–19. doi:10.1016/j.neuron.2009.11.031. PMC 2822727. PMID 20152109.
- Finger, S (2001). Origins of Neuroscience: A History of Explorations Into Brain Function [Nguồn gốc của khoa học thần kinh: Lịch sử khám phá chức năng não]. Oxford University Press US. ISBN 978-0-19-514694-3.
- Garcia-Segura LM (2009). Hormones and Brain Plasticity (Hormone và sự mềm dẻo của não). Oxford University Press US. ISBN 978-0-19-532661-1.
- Woon FL, Sood S, Hedges DW (tháng 10 năm 2010). “Hippocampal volume deficits associated with exposure to psychological trauma and posttraumatic stress disorder in adults: a meta-analysis” [Giảm thể tích vùng hải mã liên quan đến chấn thương tâm lý và rối loạn căng thẳng sau chấn thương ở người lớn: phân tích tổng hợp]. Progress in Neuro-Psychopharmacology & Biological Psychiatry. 34 (7): 1181–1188. doi:10.1016/j.pnpbp.2010.06.016. PMID 20600466.
- Gorwood P, Corruble E, Falissard B, Goodwin GM (tháng 6 năm 2008). “Toxic effects of depression on brain function: impairment of delayed recall and the cumulative length of depressive disorder in a large sample of depressed outpatients (Trầm cảm gây ảnh hưởng xấu lên chức năng não: suy giảm khả năng hồi phục và thời gian tích lũy trên nghiên cứu lấy mẫu bệnh nhân trầm cảm điều trị ngoại trú)”. The American Journal of Psychiatry. 165 (6): 731–9. doi:10.1176/appi.ajp.2008.07040574. PMID 18381906.
- Goto Y, Grace AA (tháng 11 năm 2008). “Limbic and cortical information processing in the nucleus accumbens” [Xử lý thông tin hệ viền và vỏ não trong nhân nằm]. Trends in Neurosciences. 31 (11): 552–8. doi:10.1016/j.tins.2008.08.002. PMC 2884964. PMID 18786735.
- Gray JA, McNaughton N (2000). The Neuropsychology of Anxiety: An Enquiry into the Functions of the Septo-Hippocampal System [Thần kinh học về lo âu: Khảo sát về chức năng của hệ vách-hải mã]. Oxford University Press.
- Gross CG (tháng 10 năm 1993). “Hippocampus minor and man's place in nature: a case study in the social construction of neuroanatomy” [Vai trò của hải mã bé đối với người trong tự nhiên: nghiên cứu xây dựng xã hội dưới quan điểm phẫu thuật thần kinh]. Hippocampus. 3 (4): 403–416. doi:10.1002/hipo.450030403. PMID 8269033.
- Hampel H, Bürger K, Teipel SJ, Bokde AL, Zetterberg H, Blennow K (tháng 1 năm 2008). “Core candidate neurochemical and imaging biomarkers of Alzheimer's disease” [Hóa học thần kinh và hình ảnh của bệnh Alzheimer]. Alzheimer's & Dementia. 4 (1): 38–48. doi:10.1016/j.jalz.2007.08.006. PMID 18631949.
- Harrison PJ (tháng 6 năm 2004). “The hippocampus in schizophrenia: a review of the neuropathological evidence and its pathophysiological implications” [Hải mã trong bệnh tâm thần phân liệt: tổng quan các bằng chứng về bệnh lý thần kinh và ý nghĩa sinh lý bệnh]. Psychopharmacology. 174 (1): 151–62. doi:10.1007/s00213-003-1761-y. PMID 15205886.
- Hebb DO (1949). Organization of Behavior: a Neuropsychological Theory [Tổ chức hành vi: Lý thuyết thần kinh học]. New York: John Wiley. ISBN 978-0-471-36727-7.
- Huerta PT, Lisman JE (tháng 8 năm 1993). “Heightened synaptic plasticity of hippocampal CA1 neurons during a cholinergically induced rhythmic state” [Độ dẻo synap tăng cao ở các tế bào thần kinh diện CA1 của hải mã trong trạng thái nhịp gây ra bởi hệ cholinergic]. Nature. 364 (6439): 723–5. Bibcode:1993Natur.364..723H. doi:10.1038/364723a0. PMID 8355787.
- Vertes RP (tháng 1 năm 2015). “Limbic circuitry of the midline thalamus” [Sợi dẫn truyền hệ viền của đồi thị giữa]. NCBI. doi:10.1016/j.neubiorev.2015.01.014. PMC 4976455. PMID 8355787.
- Jackson JC, Johnson A, Redish AD (tháng 11 năm 2006). “Hippocampal sharp waves and reactivation during awake states depend on repeated sequential experience” [Sóng nhọn hải mã và sự tái kích hoạt trong trạng thái tỉnh táo phụ thuộc trải nghiệm tuần tự lặp đi lặp lại]. The Journal of Neuroscience. 26 (48): 12415–26. doi:10.1523/JNEUROSCI.4118-06.2006. PMC 6674885. PMID 17135403.
- Jacobs LF, Gaulin SJ, Sherry DF, Hoffman GE (tháng 8 năm 1990). “Evolution of spatial cognition: sex-specific patterns of spatial behavior predict hippocampal size” [Tiến hóa nhận thức không gian: mô hình đặc trưng giới tính của hành vi không gian giúp dự đoán kích thước hải mã]. Proceedings of the National Academy of Sciences of the United States of America. 87 (16): 6349–52. Bibcode:1990PNAS...87.6349J. doi:10.1073/pnas.87.16.6349. PMC 54531. PMID 2201026.
- Jacobs LF (2003). “The evolution of the cognitive map” [Sự tiến hóa của bản đồ nhận thức]. Brain, Behavior and Evolution. 62 (2): 128–39. doi:10.1159/000072443. PMID 12937351.
- Jung MW, Wiener SI, McNaughton BL (tháng 12 năm 1994). “Comparison of spatial firing characteristics of units in dorsal and ventral hippocampus of the rat” [So sánh các đặc điểm phóng điện thế đối với nhận thức không gian của các cấu trúc vùng sau và vùng trước hải mã của chuột]. The Journal of Neuroscience. 14 (12): 7347–7356. doi:10.1523/JNEUROSCI.14-12-07347.1994. PMC 6576902. PMID 7996180.
- Kahana MJ, Seelig D, Madsen JR (tháng 12 năm 2001). “Theta returns” [Sóng theta dội lại]. Current Opinion in Neurobiology. 11 (6): 739–44. doi:10.1016/S0959-4388(01)00278-1. PMID 11741027.
- Kandel, Eric R.; Schwartz, James H.; Jessell, Thomas M.; Siegelbaum, Steven A.; Hudspeth, A. J. (2012). Principles of Neural Science [Nguyên lý khoa học thần kinh] (ấn bản 5). New York: McGraw-Hill Medical. tr. 1490–1491. ISBN 9780071390118. OCLC 820110349.
- Kötter R, Stephan KE (1997). “Useless or helpful? The "limbic system" concept (Vô dụng hay hữu ích? Nội dung về "hệ viền")”. Reviews in the Neurosciences. 8 (2): 139–45. doi:10.1515/REVNEURO.1997.8.2.139. PMID 9344183.
- Joëls M (tháng 4 năm 2008). “Functional actions of corticosteroids in the hippocampus (Vai trò của corticosteroid đối với hải mã)”. European Journal of Pharmacology. 583 (2–3): 312–321. doi:10.1016/j.ejphar.2007.11.064. PMID 18275953.
- Kuruba R, Hattiangady B, Shetty AK (tháng 1 năm 2009). “Hippocampal neurogenesis and neural stem cells in temporal lobe epilepsy” [Thần kinh hải mã và vai trò tế bào gốc thần kinh trong bệnh động kinh thùy thái dương]. Epilepsy & Behavior. 14 Suppl 1: 65–73. doi:10.1016/j.yebeh.2008.08.020. PMC 2654382. PMID 18796338.
- Lubenov EV, Siapas AG (tháng 5 năm 2009). “Hippocampal theta oscillations are travelling waves” [Dao động sóng theta hải mã là sóng lan truyền] (PDF). Nature. 459 (7246): 534–9. Bibcode:2009Natur.459..534L. doi:10.1038/nature08010. PMID 19489117.
- Maguire EA, Burgess N, Donnett JG, Frackowiak RS, Frith CD, O'Keefe J (tháng 5 năm 1998). “Knowing where and getting there: a human navigation network” [Nhận biết và đi đến địa điểm cho sẵn: mạng lưới điều hướng của loài người]. Science. 280 (5365): 921–24. Bibcode:1998Sci...280..921M. doi:10.1126/science.280.5365.921. PMID 9572740.
- Maguire EA, Gadian DG, Johnsrude IS, Good CD, Ashburner J, Frackowiak RS, Frith CD (tháng 4 năm 2000). “Navigation-related structural change in the hippocampi of taxi drivers” [Thay đổi cấu trúc liên quan đến điều hướng trong vùng hải mã của các tài xế taxi]. Proceedings of the National Academy of Sciences of the United States of America. 97 (8): 4398–403. Bibcode:2000PNAS...97.4398M. doi:10.1073/pnas.070039597. PMC 18253. PMID 10716738.
- Malenka RC, Bear MF (tháng 9 năm 2004). “LTP and LTD: an embarrassment of riches” [Điện thế hóa dài hạn và trầm cảm dài hạn]. Neuron. 44 (1): 5–21. doi:10.1016/j.neuron.2004.09.012. PMID 15450156.
- Matsumura N, Nishijo H, Tamura R, Eifuku S, Endo S, Ono T (tháng 3 năm 1999). “Spatial- and task-dependent neuronal responses during real and virtual translocation in the monkey hippocampal formation” [Phản ứng thần kinh phụ thuộc vào không gian và nhiệm vụ đối với quá trình hình thành hải mã khỉ]. The Journal of Neuroscience. 19 (6): 2381–93. doi:10.1523/JNEUROSCI.19-06-02381.1999. PMC 6782547. PMID 10066288.
- McNaughton BL, Battaglia FP, Jensen O, Moser EI, Moser MB (tháng 8 năm 2006). “Path integration and the neural basis of the 'cognitive map'” [Tích hợp đường đi và cơ sở thần kinh của 'bản đồ nhận thức']. Nature Reviews. Neuroscience. 7 (8): 663–78. doi:10.1038/nrn1932. PMID 16858394.
- Mizunami M, Weibrecht JM, Strausfeld NJ (tháng 12 năm 1998). “Mushroom bodies of the cockroach: their participation in place memory” [Thể cuống của gián: vai trò trí nhớ vị trí]. The Journal of Comparative Neurology. 402 (4): 520–37. doi:10.1002/(SICI)1096-9861(19981228)402:4<520::AID-CNE6>3.0.CO;2-K. PMID 9862324.
- Morris RG, Garrud P, Rawlins JN, O'Keefe J (tháng 6 năm 1982). “Place navigation impaired in rats with hippocampal lesions” [Điều hướng vị trí bị suy yếu ở chuột bị tổn thương hải mã]. Nature. 297 (5868): 681–83. Bibcode:1982Natur.297..681M. doi:10.1038/297681a0. PMID 7088155.
- Moser MB, Moser EI (1998). “Functional differentiation in the hippocampus” [Phân biệt chức năng ở vùng hải mã]. Hippocampus. 8 (6): 608–19. doi:10.1002/(SICI)1098-1063(1998)8:6<608::AID-HIPO3>3.0.CO;2-7. PMID 9882018.
- Moser EI, Kropff E, Moser MB (2008). “Place cells, grid cells, and the brain's spatial representation system” [Tế bào chỗ, tế bào lưới và hệ thống biểu diễn không gian của não]. Annual Review of Neuroscience. 31: 69–89. doi:10.1146/annurev.neuro.31.061307.090723. PMID 18284371.
- Nadel L, O'Keefe J, Black A (tháng 6 năm 1975). “Slam on the brakes: a critique of Altman, Brunner, and Bayer's response-inhibition model of hippocampal function” [Bài phê bình về mô hình của Altman, Brunner và Baye đối với chức năng hồi hải mã trong việc ức chế phản ứng]. Behavioral Biology. 14 (2): 151–62. doi:10.1016/S0091-6773(75)90148-0. PMID 1137539.
- Nakazawa K, McHugh TJ, Wilson MA, Tonegawa S (tháng 5 năm 2004). “NMDA receptors, place cells and hippocampal spatial memory” [Thụ thể NMDA, tế bào chỗ và trí nhớ không gian hồi hải mã]. Nature Reviews. Neuroscience. 5 (5): 361–72. doi:10.1038/nrn1385. PMID 15100719.
- Nieuwenhuys, R (1982). “An Overview of the Organization of the Brain of Actinopterygian Fishes (Tổng quan về tổ chức bộ não cá Actinoppetgian)”. Am. Zool. 22 (2): 287–310. doi:10.1093/icb/22.2.287.
- Numan (1995). “Effects of medial septal lesions on an operant go/no-go delayed response alternation task in rats” [Ảnh hưởng của tổn thương vùng vách trong đối với tác nhân thay thế đáp ứng chậm đi (hay không đi) ở chuột.]. Physiology & Behavior. 58 (6): 1263–1271. doi:10.1016/0031-9384(95)02044-6. PMID 8623030.
- O'Kane G, Kensinger EA, Corkin S (2004). “Evidence for semantic learning in profound amnesia: an investigation with patient H.M” [Bằng chứng về khả năng học ngữ nghĩa trong chứng mất trí nhớ sâu: thăm dò bệnh nhân H.M]. Hippocampus. 14 (4): 417–25. doi:10.1002/hipo.20005. PMID 15224979.
- O'Keefe J, Dostrovsky J (tháng 11 năm 1971). “The hippocampus as a spatial map. Preliminary evidence from unit activity in the freely-moving rat” [Hải mã như một bản đồ không gian. Bằng chứng sơ bộ về đơn vị hoạt động ở chuột khi di chuyển tự do]. Brain Research. 34 (1): 171–75. doi:10.1016/0006-8993(71)90358-1. PMID 5124915.
- O'Keefe J, Nadel L (1978). The Hippocampus as a Cognitive Map [Hải mã như một bản đồ nhận thức]. Oxford University Press. Bản gốc lưu trữ ngày 24 tháng 3 năm 2011. Truy cập ngày 18 tháng 3 năm 2020.
- Portavella M, Vargas JP, Torres B, Salas C (2002). “The effects of telencephalic pallial lesions on spatial, temporal, and emotional learning in goldfish” [Những ảnh hưởng của tổn thương vỏ đại não đối với học tập không gian, nhận thức thời gian và cảm xúc ở cá vàng]. Brain Research Bulletin. 57 (3–4): 397–99. doi:10.1016/S0361-9230(01)00699-2. PMID 11922997.
- Pearce JM (tháng 9 năm 2001). “Ammon's horn and the hippocampus” [Sừng Amon và hải mã]. Journal of Neurology, Neurosurgery, and Psychiatry. 71 (3): 351. doi:10.1136/jnnp.71.3.351. PMC 1737533. PMID 11511709.
- Pothuizen HH, Zhang WN, Jongen-Rêlo AL, Feldon J, Yee BK (tháng 2 năm 2004). “Dissociation of function between the dorsal and the ventral hippocampus in spatial learning abilities of the rat: a within-subject, within-task comparison of reference and working spatial memory” [Sự phân chia chức năng giữa vùng sau và vùng trước hải mã trong khả năng học tập không gian của chuột]. The European Journal of Neuroscience. 19 (3): 705–712. doi:10.1111/j.0953-816X.2004.03170.x. PMID 14984421.
- Prull MW, Gabrieli JD, Bunge SA (2000). “Ch 2. Age-related changes in memory: A cognitive neuroscience perspective”. Trong Craik FI, Salthouse TA (biên tập). The handbook of aging and cognition [Cẩm nang về lão hóa và nhận thức]. Erlbaum. ISBN 978-0-8058-2966-2.
- Rodríguez F, López JC, Vargas JP, Broglio C, Gómez Y, Salas C (2002). “Spatial memory and hippocampal pallium through vertebrate evolution: insights from reptiles and teleost fish” [Trí nhớ không gian và áo não hải mã thông qua quá trình tiến hóa của động vật có xương sống: những hiểu biết sâu về loài bò sát và cá xương thật]. Brain Research Bulletin. 57 (3–4): 499–503. doi:10.1016/S0361-9230(01)00682-7. PMID 11923018.
- Rolls ET, Xiang JZ (2006). “Spatial view cells in the primate hippocampus and memory recall” [Các tế bào mang vai trò không gian của hải mã linh trưởng và trí nhớ hồi tưởng]. Reviews in the Neurosciences. 17 (1–2): 175–200. doi:10.1515/REVNEURO.2006.17.1-2.175. PMID 16703951.
- Rosenzweig ES, Barnes CA (tháng 2 năm 2003). “Impact of aging on hippocampal function: plasticity, network dynamics, and cognition” [Tác động của lão hóa đến chức năng hải mã: độ dẻo, động lực học mạng lưới và nhận thức]. Progress in Neurobiology. 69 (3): 143–79. doi:10.1016/S0301-0082(02)00126-0. PMID 12758108.
- Scoville WB, Milner B (tháng 2 năm 1957). “Loss of recent memory after bilateral hippocampal lesions” [Mất trí nhớ ngắn hạn sau khi tổn thương hải mã hai bên]. Journal of Neurology, Neurosurgery, and Psychiatry. 20 (1): 11–21. doi:10.1136/jnnp.20.1.11. PMC 497229. PMID 13406589.
- Shettleworth SJ (2003). “Memory and hippocampal specialization in food-storing birds: challenges for research on comparative cognition” [Trí nhớ và vai trò của vùng đồi thị ở chim khi trữ thức ăn: những thách thức đối với nghiên cứu nhận thức so sánh]. Brain, Behavior and Evolution. 62 (2): 108–16. doi:10.1159/000072441. PMID 12937349.
- Skaggs WE, McNaughton BL, Wilson MA, Barnes CA (1996). “Theta phase precession in hippocampal neuronal populations and the compression of temporal sequences” [Suy đoán pha theta trong quần thể tế bào thần kinh hải mã và thùy thái dương]. Hippocampus. 6 (2): 149–76. doi:10.1002/(SICI)1098-1063(1996)6:2<149::AID-HIPO6>3.0.CO;2-K. PMID 8797016.
- Skaggs WE, McNaughton BL, Permenter M, Archibeque M, Vogt J, Amaral DG, Barnes CA (tháng 8 năm 2007). “EEG sharp waves and sparse ensemble unit activity in the macaque hippocampus” [Sóng nhọn trên bản điện não đồ và hoạt động đồng bộ của hải mã khỉ macaque]. Journal of Neurophysiology. 98 (2): 898–910. doi:10.1152/jn.00401.2007. PMID 17522177.
- Sloviter RS (tháng 2 năm 2005). “The neurobiology of temporal lobe epilepsy: too much information, not enough knowledge” [Sinh học thần kinh bệnh động kinh thùy thái dương: quá nhiều thông tin nhưng không đủ kiến thức]. Comptes Rendus Biologies. 328 (2): 143–53. doi:10.1016/j.crvi.2004.10.010. PMID 15771000.
- Smith DM, Mizumori SJ (2006). “Hippocampal place cells, context, and episodic memory” [Tế bào chỗ hải mã, trí nhớ bối cảnh và trí nhớ tình tiết]. Hippocampus. 16 (9): 716–29. doi:10.1002/hipo.20208. PMID 16897724.
- Solstad T, Boccara CN, Kropff E, Moser MB, Moser EI (tháng 12 năm 2008). “Representation of geometric borders in the entorhinal cortex” [Biểu diễn hình học vỏ não]. Science. 322 (5909): 1865–68. Bibcode:2008Sci...322.1865S. doi:10.1126/science.1166466. PMID 19095945.
- Squire LR (tháng 4 năm 1992). “Memory and the hippocampus: a synthesis from findings with rats, monkeys, and humans” [Trí nhớ trong mối tương quan với hải mã: tổng hợp từ những phát hiện với chuột, khỉ và người]. Psychological Review. 99 (2): 195–231. doi:10.1037/0033-295X.99.2.195. PMID 1594723.
- Squire LR, Schacter DL (2002). The Neuropsychology of Memory [Thần kinh học trí nhớ]. Guilford Press.
- Squire LR (tháng 1 năm 2009). “The legacy of patient H.M. for neuroscience” [Di sản của bệnh nhân H.M. với ngành khoa học thần kinh]. Neuron. 61 (1): 6–9. doi:10.1016/j.neuron.2008.12.023. PMC 2649674. PMID 19146808.
- Sutherland GR, McNaughton B (tháng 4 năm 2000). “Memory trace reactivation in hippocampal and neocortical neuronal ensembles” [Tái kích hoạt dấu vết ký ức trong quần thể nơron vùng hải mã và vùng vỏ não]. Current Opinion in Neurobiology. 10 (2): 180–86. doi:10.1016/S0959-4388(00)00079-9. PMID 10753801.
- Sutherland RJ, Kolb B, Whishaw IQ (tháng 8 năm 1982). “Spatial mapping: definitive disruption by hippocampal or medial frontal cortical damage in the rat” [Lập bản đồ không gian: sự gián đoạn do tổn thương hải mã và vỏ não trán trong ở chuột]. Neuroscience Letters. 31 (3): 271–6. doi:10.1016/0304-3940(82)90032-5. PMID 7133562.
- Sutherland RJ, Weisend MP, Mumby D, Astur RS, Hanlon FM, Koerner A, Thomas MJ, Wu Y, Moses SN, Cole C, Hamilton DA, Hoesing JM (2001). “Retrograde amnesia after hippocampal damage: recent vs. remote memories in two tasks” [Mất trí nhớ hồi phục sau tổn thương hồi hải mã: trí nhớ ngắn hạn và trí nhớ dài hạn]. Hippocampus. 11 (1): 27–42. doi:10.1002/1098-1063(2001)11:1<27::AID-HIPO1017>3.0.CO;2-4. PMID 11261770.
- Suzuki M, Hagino H, Nohara S, Zhou SY, Kawasaki Y, Takahashi T, Matsui M, Seto H, Ono T, Kurachi M (tháng 2 năm 2005). “Male-specific volume expansion of the human hippocampus during adolescence” [Sự tăng kích thước của hải mã ở người trong tuổi thiếu niên]. Cerebral Cortex. 15 (2): 187–93. doi:10.1093/cercor/bhh121. PMID 15238436.
- Vanderwolf CH (tháng 12 năm 2001). “The hippocampus as an olfacto-motor mechanism: were the classical anatomists right after all?” [Hải mã có cơ chế vận động khứu giác: rốt cuộc các nhà giải phẫu cổ điển có đúng?]. Behavioural Brain Research. 127 (1–2): 25–47. doi:10.1016/S0166-4328(01)00354-0. PMID 11718883.
- Vargas JP, Bingman VP, Portavella M, López JC (tháng 11 năm 2006). “Telencephalon and geometric space in goldfish” [Đại não của cá vàng]. The European Journal of Neuroscience. 24 (10): 2870–78. doi:10.1111/j.1460-9568.2006.05174.x. PMID 17156211.
- VanElzakker M, Fevurly RD, Breindel T, Spencer RL (tháng 12 năm 2008). “Environmental novelty is associated with a selective increase in Fos expression in the output elements of the hippocampal formation and the perirhinal cortex” [Sự thay cũ đổi mới của môi trường có liên quan đến sự gia tăng có chọn lọc biểu hiện Fos trong sợi ra của hải mã và vỏ não cận khứu]. Learning & Memory. 15 (12): 899–908. doi:10.1101/lm.1196508. PMC 2632843. PMID 19050162.
- Wechsler RT, Morss, AM, Wustoff, CJ, & Caughey, AB (2004). Blueprints notes & cases: Neuroscience [Ghi chú & ca lâm sàng: Khoa học thần kinh]. Oxford: Blackwell Publishing. tr. 37. ISBN 978-1-4051-0349-7.Quản lý CS1: nhiều tên: danh sách tác giả (liên kết)
- West MJ (1990). “Chapter 2 Stereological studies of the hippocampus: A comparison of the hippocampal subdivisions of diverse species including hedgehogs, laboratory rodents, wild mice and men”. Stereological studies of the hippocampus: a comparison of the hippocampal subdivisions of diverse species including hedgehogs, laboratory rodents, wild mice and men [Các nghiên cứu lập thể về hải mã: so sánh các phân vùng hải mã của các loài khác nhau: nhím, chuột thí nghiệm, chuột hoang và người]. Progress in Brain Research. 83. tr. 13–36. doi:10.1016/S0079-6123(08)61238-8. ISBN 9780444811493. PMID 2203095.
- Wilson MA, McNaughton BL (tháng 7 năm 1994). “Reactivation of hippocampal ensemble memories during sleep” [Tái kích hoạt ký ức ở quần thể nơron hải mã trong khi ngủ]. Science. 265 (5172): 676–79. Bibcode:1994Sci...265..676W. doi:10.1126/science.8036517. PMID 8036517.
- Winson J (tháng 7 năm 1978). “Loss of hippocampal theta rhythm results in spatial memory deficit in the rat” [Mất nhịp theta hải mã dẫn đến suy giảm nhớ không gian ở chuột]. Science. 201 (4351): 160–63. Bibcode:1978Sci...201..160W. doi:10.1126/science.663646. PMID 663646.
- Docampo-Seara A, Lagadec R, Mazan S, Rodríguez MA, Quintana-Urzainqui I, Candal E (tháng 7 năm 2018). “Study of pallial neurogenesis in shark embryos and the evolutionary origin of the subventricular zone”. Brain Structure and Function. 223 (8): 3593–3612. doi:10.1007/s00429-018-1705-2. PMID 29980930.
- Sách
- Trịnh Văn Minh (2017). Giải phẫu người (Tập 3: Hệ thần kinh - Hệ nội tiết). Nhà xuất bản Giáo dục Việt Nam. ISBN 978-604-0-04586-7.
Liên kết ngoài
- Hippocampus tại Encyclopædia Britannica (tiếng Anh)
- Hình ảnh lát não nhuộm màu bao gồm "hippocampus" tại dự án BrainMaps
- Diagram of a Hippocampal Brain Slice
- Hippocampus – Cell Centered Database
- Temporal-lobe.com An interactive diagram of the rat parahippocampal-hippocampal region
- Search Hippocampus on BrainNavigator via BrainNavigator
- Buzsaki G (2010). “Hippocampus”. Scholarpedia. 6 (1): 1468. Bibcode:2011SchpJ...6.1468B. doi:10.4249/scholarpedia.1468.
- Docampo-Seara A, Lagadec R, Mazan S, Rodríguez MA, Quintana-Urzainqui I, Candal E (tháng 7 năm 2018). “Study of pallial neurogenesis in shark embryos and the evolutionary origin of the subventricular zone”. Brain Structure and Function. 223 (8): 3593–3612. doi:10.1007/s00429-018-1705-2. PMID 29980930.
- Frank H.Netter, MD (2017). Atlas Giải phẫu người, Vietnamese Edition (ấn bản 6). Nhà xuất bản Y học, ELSEVIER. ISBN 978-604-66-1320-6.
- Frank H.Netter, MD (2017). Atlas of Human Anatomy (ấn bản 7). ELSEVIER MASSON. ISBN 978-604-66-1320-6.
- Frank H.Netter, MD (2017). Atlas d'anatomie humaine (ấn bản 5). ELSEVIER MASSON. ISBN 978-229-47-1297-5.
- Bài giảng Giải phẫu học, PGS Nguyễn Quang Quyền, tái bản lần thứ mười lăm
- PGS.TS Nguyễn Quang Huy (2017). Giải phẫu người (ấn bản 2). Nhà xuất bản Y học. ISBN 978-604-66-2933-7.
- Phiên bản trực tuyến sách Gray's Anatomy — Giải phẫu cơ thể người, Gray, tái bản lần thứ hai mươi (năm 1918).
- Gray's Anatomy, tái bản lần thứ nhất, năm 1858 (liên kết đến file PDF)