

Tiến trình tiến hóa của động vật có vú đã trải qua nhiều giai đoạn kể từ khi tổ tiên của chúng, động vật Một cung bên, lần đầu tiên xuất hiện vào cuối kỷ Than đá. Vào giữa kỷ Tam Điệp, nhiều loài Một cung bên trông giống động vật có vú bắt đầu xuất hiện. Dòng dõi dẫn đến động vật có vú ngày nay rẽ nhánh vào kỷ Jura; thú Một cung bên từ thời kỳ này bao gồm chi Dryolestes, quan hệ gần hơn với các thú có nhau thai (placentalia) và thú có túi (marsupial) hiện đại hơn là các loài thú thuộc bộ Đơn huyệt (monotreme), các chi khác như Ambondro, thì quan hệ gần hơn với thú đơn huyệt. Sau này, hai dòng dõi eutherian và metatherian tách ra; metatherian là những động vật có họ hàng gần hơn với thú có túi còn eutherian gần gũi hơn với thú có nhau thai. Vì Juramaia (loài thú eutheria được biết đến sớm nhất) sống cách đây khoảng 160 triệu năm trong kỷ Jura, sự tách rời giữa hai dòng thú này chắc hẳn đã phải xảy ra trong cùng thời kỳ đó.
Sau sự kiện tuyệt chủng kỷ Phấn trắng – Paleogen quét sạch những loài khủng long không phải là chim (chim là nhánh khủng long duy nhất còn sót lại) và một số nhóm động vật có vú, thú có nhau thai và thú có túi tiến hóa thành nhiều dạng mới và chiếm các hốc sinh thái bị bỏ trống trong suốt đại Tân Sinh.
Động vật có vú là nhóm động vật Một cung bên duy nhất còn tồn tại. Dòng dõi Một cung bên tách ra khỏi lớp Mặt thằn lằn vào giai đoạn cuối kỷ Than đá, khoảng 320-315 triệu năm trước. Lớp Mặt thằn lằn bao gồm các loài bò sát và chim ngày nay, cùng với tất cả các loài động vật tuyệt chủng có quan hệ mật thiết với chúng. Nhóm này không bao gồm các loài bò sát giống động vật có vú, một nhóm gần gũi hơn với các động vật có vú.
Trong suốt kỷ Permi, các động vật Một cung bên bao gồm nhiều loài ăn thịt chiếm ưu thế và một số động vật ăn cỏ quan trọng. Tuy nhiên, trong kỷ Tam Điệp kế tiếp, một nhóm Mặt thằn lằn khá khiêm tốn trước đó gọi là lớp thằn lằn chúa, đã vươn lên và chiếm ưu thế. Các động vật có vú xuất hiện trong thời kỳ này; với khứu giác vượt trội và bộ não lớn, đã tạo tiền đề cơ bản cho các thích ứng phù hợp tại hốc sinh thái ban đêm, thời điểm mà chúng ít tiếp xúc với các loài săn mồi (chủ yếu là thằn lằn chúa) lớn hơn. Lối sống về đêm có lẽ đã góp phần rất lớn vào sự phát sinh các đặc điểm của lớp thú ngày nay như lông mao giữ ấm cơ thể và hệ trao đổi chất hằng nhiệt. Vào kỷ Phấn Trắng, sau khi khủng long chân thú thay thế các loài Rauisuchia (họ hàng với cá sấu) để trở thành động vật ăn thịt chiếm ưu thế, động vật có vú phải chạy lan sang các hốc sinh thái khác. Ví dụ, một số trở thành thủy sinh, một vài trở thành loài bay lượn và một số thậm chí ăn thịt cả khủng long chưa trưởng thành.
Hầu hết các bằng chứng được lấy từ bản ghi hóa thạch. Trong quá khứ, hóa thạch thú và tổ tiên của chúng vào đại Trung sinh rất hiếm hoi và rời rạc; nhưng, từ giữa những năm 1990, đã có nhiều phát hiện mới quan trọng, đặc biệt là ở Trung Quốc. Các kỹ thuật tân tiến trong phương pháp phát sinh chủng loại phân tử (ngành nghiên cứu và ước tính thời gian mà các điểm quan trọng bắt đầu tách ra đối với các loài hiện đại) đã làm sáng tỏ một số khía cạnh của quá trình tiến hóa lớp thú. Nếu được sử dụng một cách khôn khéo, những kỹ thuật này thường xuyên đồng bộ với bản ghi hóa thạch.
Mặc dù các tuyến vú là một đặc trưng của thú hiện đại, ta không hiểu rõ sự tiến hóa của nó do các mô mềm thường không được bảo quản trong các hóa thạch. Hầu hết các nghiên cứu về sự tiến hóa của thú chỉ tập trung vào hình dạng của răng, phần cứng nhất trên cơ thể của các loài động vật bốn chi. Các đặc điểm nghiên cứu quan trọng khác bao gồm: sự tiến hóa của xương tai giữa, tư thế chân tay duỗi thẳng, vòm miệng xương thứ cấp, lông mao, tóc và hệ trao đổi chất máu nóng.
Định nghĩa
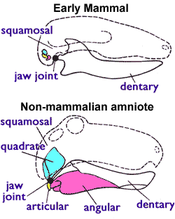
Con cái của các lớp thú còn sống sở hữu các tuyến vú sản xuất sữa và ta có thể xác định một con vật là thú nhờ đặc điểm đó. Nhưng khi nghiên cứu hóa thạch, các nhà cổ sinh vật học không thể dựa vào các tuyến vú và mô mềm để xác định một hóa thạch có phải là thú hay không, bởi vì những thứ đó hiếm khi được bảo tồn nguyên vẹn.
Thay vào đó, các nhà cổ sinh vật học dựa vào một đặc điểm khác, hiện hữu ở tất cả các động vật có vú còn sống (bao gồm cả bộ Đơn huyệt) nhưng không hiện hữu ở bộ Cung thú Tam Điệp nguyên thủy (minh họa ở Hình 1), đó chính là: động vật có vú sử dụng hai xương nhỏ ở sọ để nghe ngóng (xương nhỏ thính giác), còn tất cả các loài động vật có màng ối khác dùng để ăn. Các động vật có màng ối buổi sớm có khớp hàm cấu thành bởi xương khớp hàm (articular bone, xương nhỏ ở phía sau của hàm dưới) và xương vuông (quadrate bone, xương nhỏ ở phía sau của hàm trên). Tất cả các Tetrapod không phải động vật có vú, bao gồm các loài lưỡng cư, rùa, thằn lằn, rắn, cá sấu, khủng long (bao hàm cả hậu duệ của chúng đó chính là các loài chim), ngư long, dực long và bộ Cung thú, đều sở hữu hệ thống xương nêu trên. Nhưng ở động vật có vú thực sự thì khớp hàm chỉ bao gồm xương hàm dưới (dentary bone hay xương mang răng) và xương vảy (squamosal bone, một loại xương sọ nhỏ). Trong kỷ Jura, xương vuông và xương khớp hàm của động vật có vú tiến hóa thành xương đe (incus bone) và xương búa (malleus bone) ghép thành tai giữa. Ngoài ra, động vật có vú còn có cặp lồi cầu xương chẩm; hai núm ở đáy hộp sọ khớp với đốt sống cổ trên cùng (đốt atlas). Các Tetrapod khác chỉ có một lồi cầu xương chẩm duy nhất.
Tổ tiên của động vật có vú
|
Pelycosaur
|
Động vật có màng ối
|
−4500 —
–
—
–
−4000 —
–
—
–
−3500 —
–
—
–
−3000 —
–
—
–
−2500 —
–
—
–
−2000 —
–
—
–
−1500 —
–
—
–
−1000 —
–
—
–
−500 —
–
—
–
0 —
|
|
|
||||||||||||||||||||||||||
Những động vật có xương sống trú ngụ hoàn toàn ở trên cạn đầu tiên là những động vật có màng ối (tức là, các động vật đẻ trứng có màng trong cho phép phôi đang phát triển có thể thở nhưng vẫn giữ độ ẩm và nước bên trong). Điều này cho phép chúng đẻ trứng trên đất liền, còn các loài lưỡng cư nói chung cần phải đẻ trứng trong môi trường nước (một số loài lưỡng cư, chẳng hạn như Pipa pipa, tiến hóa theo hướng khác để vượt qua trở ngại này). Các động vật có màng ối đầu tiên dường như xuất hiện vào giữa kỷ Than đá, phái sinh từ tổ tiên reptiliomorpha (động vật dạng bò sát).
Trong vòng vài triệu năm, hai dòng dõi động vật có màng ối quan trọng nhất đó là: Lớp Một cung bên, tức tổ tiên của động vật có vú; và Lớp Mặt thằn lằn, tức tổ tiên của thằn lằn, rắn, rùa, cá sấu, khủng long và chim; tách ra khỏi nhau. Các hóa thạch sớm nhất của hai nhóm này (ví dụ Archaeothyris và Hylonomus) có niên đại vào khoảng 320-315 triệu năm trước. Thời điểm xuất hiện rất khó căn cứ, vì hóa thạch động vật có xương sống từ kì Than Đá Muộn rất hiếm, và do đó, sự xuất hiện của chúng có lẽ còn sớm hơn bản ghi hóa thạch mà ta hiện biết.
Động vật Một cung bên

Hộp sọ của động vật Một cung bên được nhận dạng bằng một đặc điểm độc đáo là các lỗ (cửa sổ) ở sau ổ mắt, thực hiện chức năng sau:
- khiến hộp sọ nhẹ đi mà không làm giảm sức mạnh của nó.
- tiết kiệm năng lượng bằng cách sử dụng ít xương hơn.
- cung cấp diện tích bám cho cơ mồm. Có các điểm bám cách xa hàm khiến cho các cơ dài hơn và do đó tạo ra lực kéo mạnh trên một phạm vi rộng của chuyển động hàm mà không bị dãn quâ dài hoặc co lại vượt quá phạm vi tối ưu của chúng.
Các pelycosaur Một cung bên bao gồm các động vật có xương sống trên cạn lớn nhất của kì Permi Sớm, chẳng hạn như Cotylorhynchus hancocki dài 6 m (20 ft). Các pelycosaur lớn khác bao gồm: Dimetrodon grandis và Edaphosaurus cruciger.
Bộ Cung thú
Bộ Cung thú (Therapsid) tiến hóa từ tổ tiên sphenacodont (phân nhánh Một cung bên sơ khai) vào giữa kỉ Permi và trở thành động vật có xương sống chiếm lĩnh trên đất liền. Hộp sọ của chúng khác động vật Một cung bên sơ khai ở một số điểm, bao gồm: cửa sổ hộp sọ rộng hơn và các răng cửa có cùng kích thước.
Dòng dõi Cung thú dẫn trực tiếp đến lớp Thú đã trải qua một loạt các giai đoạn phát triển, khởi đầu với những sinh vật rất giống tổ tiên pelycosaur và kết thúc với một số loài có thể dễ bị nhầm lẫn là động vật có vú:
- Một vòm miệng có lớp xương thứ cấp dần dần tiến hóa. Hầu hết các sách báo thường khẳng định đây là điều kiện tiên quyết cho sự tiến hóa của sự trao đổi chất cực kỳ hiệu quả ở động vật có vú bởi vì đặc điểm này cho phép chúng vừa ăn vừa thở cùng một lúc. Tuy vậy, một số nhà khoa học chỉ ra rằng nhiều loài động vật ngoại nhiệt hiện đại cũng sử dụng một vòm miệng thịt thứ cấp để tách miệng ra khỏi khí quản và rằng vòm miệng xương cung cấp một bề mặt mà lưỡi có thể trộn thức ăn, tạo điều kiện cho hành động nhai thay vì thở. Cách giải thích vòm miệng xương thứ cấp như sự hỗ trợ cho hành động nhai cũng thể hiện sự tiến bộ của quá trình trao đổi chất nhanh hơn, bởi vì nhai thức ăn giúp làm giảm kích thước của các hạt dinh dưỡng rồi được đưa vào dạ dày và do đó tăng tốc độ tiêu hóa. Ở động vật có vú, vòm miệng được hình thành bởi hai xương cụ thể, nhưng các loài Cung thú kỷ Permi khác sở hữu một kiểu kết hợp xương khác có chức năng giống hệt vòm miệng.
- Xương răng dần dần trở thành xương chính của hàm dưới.
- Tư thế chân tay duỗi thằng dần được tiến hóa, giúp tăng sức bền bỉ của chúng và tránh điều kiện biên Carrier. Quá trình này tuy vậy diễn ra thất thường và rất chậm - ví dụ: tất cả các Cung thú ăn cỏ đều có tư thế chân tay xoạc ngang (một số dạng muộn hơn có lẽ có chân sau nửa duỗi thẳng); Các Cung thú ăn thịt kỷ Permi có chân trước xoạc ngang và một số loài vào kì Permi muộn vẫn còn những chi nửa xoạc ngang. Trên thực tế, các thú đơn huyệt hiện đại vẫn có tứ chi nửa xoạc ngang.
Phát sinh loài của bộ Cung thú
Lược đồ sau đây biểu thị cây phát sinh loài dẫn đến động vật có vú hiện đại, lấy từ trang web Palaeos.com.
| Bộ Cung thú |
|
||||||||||||||||||||||||||||||||||||||||||
Chỉ có dòng dicynodont, therocephalian, và cynodont sống sót tới tận kỷ Tam Điệp.
Phân bộ Biarmosuchia
Phân bộ Biarmosuchia là nhánh nguyên thủy và giống pelycosaur nhất trong số các Cung thú.
Phân bộ Dinocephalia
Phân bộ Dinocephalia ("đầu khủng khiếp") bao gồm nhiều loài ăn thịt lẫn ăn cỏ. Chúng rất to lớn; chẳng hạn như chi Anteosaurus dài tới 6 m. Một số loài ăn thịt Dinocephalia có phần chi sau nửa duỗi thẳng, nhưng tất cả Dinocephalia đều có chi trước dạng ngang. Tựu trung, đây là phân bộ Cung thú còn khá sơ khai do chúng vẫn chưa có vòm miệng xương thứ cấp và phần hàm vẫn rất "bò sát".
Phân bộ Anomodontia

Phân bộ Anomodontia ("răng dị thường") là một trong những Phân bộ Cung thú ăn cỏ thành công nhất - một nhóm phụ của chúng đặt tên là dicynodontia, đã sống sót đến gần cuối kỉ Tam Điệp. Tuy vậy, phân bộ này rất khác so với động vật có vú ăn cỏ hiện đại, vì chúng chỉ sở hữu duy nhất cặp răng nanh ở hàm trên. Có đề xuất rằng chúng sở hữu mỏ sừng giống như ở chim hoặc các loài khủng long sừng.
Phân bộ Theriodontia
Phân bộ Bò sát răng thú (Theriodontia) và hậu duệ của chúng có xương khớp hàm dưới gắn chặt với xương vuông nhỏ của hộp sọ. Điều này cho phép chúng há mồm rộng hơn; ví dụ các loài gorgonopsian ("mặt gorgon") ăn thịt tiến hóa thêm "răng kiếm" nhờ vào góc há mồm rất rộng của chúng. Bản lề hàm của theriodont mang tầm quan trọng lâu dài hơn - kích thước giảm của xương vuông là một bước tiến quan trọng đối với sự tiến hóa của khớp hàm và tai giữa ở động vật có vú hiện đại.
Các gorgonopsian tuy vậy vẫn bảo lưu một số đặc điểm sơ khai: không có vòm miệng xương thứ cấp (các xương khác thực hiện chức năng này); chân trước vẫn dạng ngang; chân sau có thể dạng ngang hoặc duỗi thẳng. Các therocephalia ("đầu thú") dường như xuất hiện vào cùng thời điểm với các gorgonopsian, sở hữu thêm các đặc điểm giống ở động vật có vú, ví dụ: xương ngón tay và ngón chân của chúng có cùng số lượng đốt ngón như ở động vật có vú sớm (và cùng số lượng như ở các loài linh trưởng, bao gồm cả con người).
Phân bộ Cynodontia

Phân bộ Một răng chó (Cynodontia), một nhóm Cung thú phái sinh vào cuối kỷ Permi, là nhánh tổ tiên của tất cả các động vật có vú hiện đại. Các đặc điểm giống động vật có vú ở Cynodont bao gồm: số lượng xương ở hàm dưới tiêu giảm, có vòm miệng xương thứ cấp, răng má có hoa văn phức tạp ở vòng đỉnh răng và bộ não nằm trong khoang nội sọ (endocranial cavity).
Các nhà khảo cổ đã tìm thấy các hệ thống hang nhiều buồng, có hang chôn vùi 20 bộ xương của chi Trirachodon với niên đại vào kỉ Tam Điệp sớm. Những con vật này có vẻ như đã chết đuối sau một trận lũ quét. Các hang này có sức chứa rất lớn, chứng minh rằng những loài vật này có khả năng thực hiện các hành vi xã hội phức tạp.
Sự chiếm lĩnh kỉ Tam Điệp
Sự kiện tuyệt chủng kỷ Permi–kỷ Trias thảm khốc đã tiêu diệt khoảng 70% loài động vật có xương sống và phần lớn các thực vật trên cạn vào khoảng hơn 250 triệu năm trước.
Kết quả là hệ sinh thái cùng các mạng lưới thức ăn sụp đổ, và phải mất khoảng 30 triệu năm thì hệ sinh thái mới được tái thiết hoàn toàn. Theo sau sự tuyệt chủng của các loài gorgonopsid (những kẻ săn mồi chiếm ưu thế ở cuối kỷ Permi) đối thủ cạnh tranh chính của cynodont để thống trị các hốc sinh thái ăn thịt là một nhóm Mặt thằn lằn khá khiêm tốn trước đó, những loài Thằn lằn chúa.
Các thằn lằn chúa nhanh chóng trở thành động vật ăn thịt chiếm ưu thế, và sự kiện này được các nhà cổ sinh gọi là "sự chiếm lĩnh kỉ Tam Điệp". Thành công của chúng có lẽ là do kỷ Tam Điệp sớm rất khô cằn và hệ thống bảo toàn nước vượt trội của thằn lằn chúa mang lại cho chúng một lợi thế mang tính quyết định. Tất cả các thằn lằn chúa được biết đều có lớp da không có tuyến và chúng loại bỏ chất thải nitơ trong người thông qua một hỗn hợp axit uric chứa ít nước, trong khi các cynodont phải bài tiết hầu hết các chất thải như vậy qua dung dịch urê (cần một lượng nước đáng kể để giữ cho urê hòa tan, do vậy các cynodont thải nhiều nước hơn trong quá trình bài tiết) giống động vật có vú ngày nay.
Tuy nhiên, học thuyết này đang bị nghi ngờ vì nó cho rằng lớp Một cung thú không tối ưu trong việc giữ nước, rằng sự suy giảm của Một cung thú trùng với sự thay đổi khí hậu hoặc sự đa dạng hóa của thằn lằn chúa (cả hai khẳng định này chưa được kiểm nghiệm) và thực tế là, động vật có vú sống ở sa mạc cũng thích nghi tốt trong môi trường khô cằn không khác mấy so với thằn lằn chúa, thậm chí một số loài cynodont như Trucidocynodon là những kẻ săn mồi có kích thước lớn.
Sự chiếm lĩnh kỉ Tam Điệp có lẽ là một sự kiện quan trọng trong quá trình tiến hóa của thú. Hai nhóm xuất phát từ phân bộ cynodont đầu tiên rất thành công ở các hốc sinh thái không cạnh tranh với các loài thằn lằn chúa, đó là: tritylodont (hầu hết ăn cỏ) và động vật có vú (hầu hết là động vật ăn côn trùng sống về đêm, mặc dù có một số chi như Sinoconodon là những kẻ săn mồi lớn). Kết quả là:
- Các Cung thú có xu hướng rất nhanh để tiến hóa nhiều loại răng đa dạng với ghép khớp chính xác, xuất phát từ nhu cầu bắt giữ các động vật chân đốt và nghiền nát vỏ của chúng.
- Kích cỡ cơ thể tổ tiên thú giảm xuống dưới 50 mm, khiến cho những tiến bộ trong lớp cách nhiệt và điều chỉnh thân nhiệt rất cần thiết cho cuộc sống về đêm.
- Chúng tiến hóa thính giác và khứu giác cực kỳ nhạy bén.
- Tai giữa phát triển
- Sự gia tăng kích thước của thùy khứu giác ở não, làm tăng trọng lượng của não theo phần trăm tổng trọng lượng cơ thể. Mô não đòi hỏi một nguồn năng lượng rất lớn. Nhu cầu cần nhiều thức ăn để hỗ trợ bộ não lại gây tiếp áp lực lên sự phát triển về cách nhiệt, điều chỉnh thân nhiệt.
- Có lẽ do tác dụng phụ của cuộc sống về đêm, động vật có vú mất đi hai trong số bốn tế bào hình nón (tế bào cảm quang trong võng mạc), đặc điểm kế thừa rất xa xưa của các động vật có màng ối. Nghịch lý thay, điều này có lẽ đã cải thiện khả năng phân biệt màu sắc trong ánh sáng mờ của thú có vú.
Sự rút lui về hốc sinh thái ban đêm còn được các nhà cổ sinh gọi là nút cổ chai về đêm, giúp lý giải nhiều đặc điểm hiện nay của động vật có vú.
Từ cynodontia đến thú có vú chỏm cây
Bản ghi hóa thạch
Động vật Một cung bên trong đại Trung sinh đã phát triển khớp hàm gồm xương răng và xương squamosal, được bảo tồn chỉ trong các mẫu hóa thạch tốt, chủ yếu là vì kích thước của chúng nhỏ hơn cả chuột:
- Chúng phần lớn chúng sinh sống ở những môi trường ít có khả năng cung cấp hóa thạch tốt. Bãi bồi là môi trường đất liền rất thuận lợi để hóa thạch hình thành, nhưng vì những nơi này bị chi phối bởi các động vật có cỡ từ trung bình đến lớn và các động vật có vú không thể cạnh tranh với các loài thằn lằn chúa có kích cỡ từ trung bình đến lớn, nên hóa thạch của động vật có vú rất hiếm. Các đường dấu chân từ kỳ Phấn trắng Sớm ở Angola chỉ ra rằng một loài thú cỡ gấu mèo đã từng tồn tại ở đây 118 triệu năm trước.
- Xương của chúng dễ bị phá hủy trước khi có thể bị hóa thạch - bởi xác thịt và xương cốt thường bị ăn bởi các loài thú khác, bị tiêu hủy bởi nấm và vi khuẩn, hoặc bị giẫm đạp.
- Hóa thạch nhỏ khó phát hiện hơn và dễ bị phong hóa hơn.
Tuy nhiên, trong 50 năm qua, số lượng hóa thạch động vật có vú Đại Trung sinh đã tăng lên đáng kể; chẳng hạn, 116 chi được biết đến vào năm 1979 và tăng lên thành khoảng 310 chi vào năm 2007. Chất lượng cũng tăng đến mức "ít nhất 18 loài thú đại Trung sinh có mẫu điển hình gần như hoàn chỉnh".
Động vật có vú và Mammaliaform (Động vật dạng động vật có vú)
Một số tác giả hạn chế sử dụng thuật ngữ "động vật có vú" đối với nhóm chỏm cây, bao gồm tổ tiên chung gần đây nhất của các loài đơn huyệt, thú có túi và thú có nhau thai, cùng với tất cả các hậu duệ của loài tổ tiên đó. Trong một bài báo có ảnh hưởng năm 1988, Timothy Rowe đã ủng hộ sự hạn chế này, lập luận rằng "tổ tiên... cung cấp phương tiện duy nhất để xác định chính xác các loài" và đặc biệt là sự tách ra của các loài đơn huyệt khỏi các động vật liên quan chặt chẽ hơn với thú có túi và nhau thai "là mối quan tâm chính đối với bất kỳ nghiên cứu nào về lớp Mammalia nói chung." Để cho thấy với một vài đơn vị có liên quan nằm ngoài nhóm chỏm cây, ông đã định nghĩa Mammaliaforme bao gồm" tổ tiên chung cuối cùng của Morganucodontidae và Mammalia [như ông đã định nghĩa nhóm đằng sau] và tất cả các hậu duệ của chúng." Bên cạnh Morganucodontidae, đơn vị phân loại mới được xác định bao gồm Docodonta và Kuehneotheriidae. Mặc dù bộ haramiyids đã được đề cập là động vật có vú từ những năm 1860, Rowe đã loại chúng ra khỏi nhóm Mammaliaformes vì nằm ngoài định nghĩa của ông, đưa chúng vào một nhánh lớn hơn, Mammaliamorpha.
Một số tác giả đã thông qua thuật ngữ này, để tránh sự hiểu lầm, rằng họ đã làm như vậy. Tuy nhiên, hầu hết các nhà cổ sinh vật học vẫn nghĩ rằng động vật có khớp xương răng - squamosal và răng hàm đặc trưng của động vật có vú hiện đại là thành viên của Mammalia.
Trong trường hợp sự mơ hồ của thuật ngữ "động vật có vú" có thể gây nhầm lẫn, bài viết này sử dụng các thuật ngữ "mammaliaform" và "động vật có vú chỏm cây".
| Cynodontia |
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

Morganucodontidae
Morganucodontidae xuất hiện lần đầu tiên vào cuối kỉ Tam Điệp, khoảng 205 triệu năm trước. Chúng là một ví dụ tuyệt vời của hóa thạch chuyển tiếp, vì chúng có cả khớp hàm dưới - squamosal và khớp xương vuông.
Docodonta

Docodonta, một trong số các loài động vật có vú kỷ Jura phổ biến nhất, được ghi nhận về sự tinh vi của răng hàm. Chúng được cho là có khuynh hướng bán thủy sinh nói chung, loài ăn cá Castorocauda ("đuôi hải ly"), sống vào giữa kỷ Jura khoảng 164 triệu năm trước và được phát hiện lần đầu tiên vào năm 2004 và được mô tả lần đầu tiên vào năm 2006, là một ví dụ tốt nhất của họ này. Castorocauda không phải là một động vật có vú thuộc nhóm chỏm cây, nhưng chi này cực kỳ quan trọng trong nghiên cứu về sự tiến hóa của thú có vú vì các hóa thạch gần như hoàn chỉnh (một thứ khá xa xỉ trong giới cổ sinh vật học) và nó phá vỡ định kiến rằng những loài thú cõ vú đại Trung sinh là "loài ăn côn trùng sống về đêm nhỏ":
- Nó lớn hơn đáng kể so với hầu hết các hóa thạch động vật có vú - số đo vào khoảng 43 cm từ mũi đến chóp đuôi dài 130 mm và có thể nặng từ 500-800 g.
- Nó cung cấp bằng chứng tuyệt đối sớm nhất về tóc và lông thú. Trước đây sớm nhất là ở loài Eomaia, một nhóm động vật có vú sống vào khoảng 125 triệu năm trước.
- Nó có sự thích nghi cho cuộc sống dưới nước bao gồm xương đuôi dẹt và tàn dư của mô mềm giữa mu bàn chân, cho thấy chúng có màng. Trước đây, động vật có vú bán thủy sản được biết đến sớm nhất là từ thế Thủy Tân, khoảng 110 triệu năm sau đó.
- Chân trước mạnh mẽ của Castorocauda trông rất phù hợp để đào hố. Đặc điểm này khiến cho nó tương tự như thú mỏ vịt, cũng có tập tính bơi và đào.
- Răng của nó có vẻ thích nghi để ăn cá: hai răng hàm đầu tiên có các núm theo đường thẳng, khiến chúng thích hợp để kẹp và cắt hơn là mài; và những răng hàm này được uốn cong về phía sau, để giúp kẹp chắc con mồi trơn trượt.
Hadrocodium
Cây phát sinh loài ở trên cho thấy Hadrocodium là một "cô dì chú bác" của động vật có vú chỏm cây. Loài vật này, xuất hiện vào khoảng 195 năm trước trong kỷ Jura sớm, thể hiện một số đặc điểm quan trọng:
- Hàm chỉ chứa một bộ xương răng và xương squamosal, và bộ hàm không chứa các xương nhỏ hơn ở phía sau, không giống như thiết kế của Cung thú.
- Trong bộ Cung thú và động vật có vú sớm, màng nhĩ có thể đã kéo dài qua một máng ở phía sau hàm dưới. Nhưng Hadrocodium không có máng như vậy, điều đó cho thấy rằng tai của chúng là một phần của hộp sọ, giống cấu trúc của động vật có vú chỏm cây - và do đó, hình dạng khớp và xương vuông đã được di chuyển vào tai giữa và trở thành xương đe và xương búa. Mặt khác, xương răng có một "vịnh" mà động vật có vú không có. Điều này cho thấy xương răng của Hadrocodium vẫn giữ được hình dạng tương tự như trong trường hợp khớp hàm và xương vuông đặt ở dưới, và do đó Hadrocodium có một tai giữa đầy đủ của động vật có vú.
Những thú có vú chỏm cây sớm nhất
Các động vật có vú thuộc nhóm chỏm cây, đôi khi được gọi là "động vật có vú thực sự", là các thú có vú còn tồn tại và họ hàng của chúng quay trở lại tổ tiên chung cuối cùng của chúng. Vì nhóm này có các thành viên còn sống, phân tích DNA có thể được áp dụng trong nỗ lực giải thích sự tiến hóa của các đặc điểm không xuất hiện trong hồ sơ hóa thạch. Nỗ lực này thường sử dụng phương pháp phát sinh chủng loại phân tử, một kỹ thuật đã trở nên phổ biến từ giữa những năm 1980.
Cây phát sinh loài
| Lớp Thú thuộc nhóm chỏm cây |
|
||||||||||||||||||||||||||||||||||||||||||
Phân biệt màu sắc
Các động vật có màng ối sớm sở hữu 4 tế bào nón võng mạc sử dụng để phân biệt màu sắc: một nhạy cảm với màu đỏ, một nhạy cảm với màu xanh lá và hai tế bào còn lại cảm nhận sự mạnh yếu của màu xanh lam. Tế bào nón cảm nhận màu xanh lá không được thừa hưởng bởi bất kỳ động vật có vú chỏm cây nào, nhưng tất cả các cá thể tự nhiên được thừa hưởng màu đỏ. Do đó, động vật có vú chỏm cây có ba tê bào cảm quang hình nón, một màu đỏ và cả hai màu xanh lam. Tất cả các hậu duệ còn lại của chúng đã mất một trong những tế bào nhạy cảm với màu xanh nhưng không phải lúc nào chúng cũng mất chung một tế bào xanh lam: thú Đơn huyệt giữ lại một tế bào nhạy cảm với màu xanh, trong khi đó, thú có túi và nhau thai vẫn giữ lại cái kia (trừ Bộ Cá voi, sau này cũng mất hẳn tế bào cảm nhận màu xanh khác). Một số thú nhau thai và thú có túi, bao gồm cả con người, sau đó đã tiến hóa một tê bào cảm quang nhạy cảm với màu xanh lá cây; giống như động vật có vú chỏm cây, do đó, tầm nhìn của chúng là ba màu.
Australosphenida và Ausktribosphenidae
Ausktribosphenidae là một nhóm được gán cho những phát hiện gây bối rối cho các nhà cổ sinh vật học:
- dường như có kết cấu răng hàm ba chỏm, chỉ biết đến ở thú có nhau thai và thú có túi.
- có niên đại vào giữa kỉ Phấn Trắng tìm thấy ở Australia, nhưng lúc bấy giờ, châu Úc đang được kết nối với châu Nam Cực và thú có vú có nhau thai có nguồn gốc ở Bắc bán cầu và bị giam giữ ở đó, cho tới khi các lục địa trôi dạt và ghép với nhau, như Bắc Mỹ nối với Nam Mỹ, Châu Á nối với Châu Phi và Châu Á với Ấn Độ.
- chỉ mới tìm thấy vài mẫu răng và hàm, không quá hữu dụng cho việc nghiên cứu.
Australosphenida là một nhóm động vật bao gồm Ausktribosphenidae và thú Đơn huyệt. Asfaltomylos (giữa đến cuối kỉ Jura, từ Patagonia) đã được hiểu là một Australosphenid cơ bản (có đặc điểm giống với, khác với hoặc thiếu hẳn những đặc điểm đặc hữu của Ausktribosphenidae và Đơn huyệt), và cho thấy sự đa dạng của chúng ở Gondwanaland (siêu lục địa nằm ở Nam bán cầu cũ).
Thú Đơn huyệt
Teinolophos, từ Úc, là chi đơn huyệt được biết đến sớm nhất. Một nghiên cứu năm 2007 (xuất bản 2008) cho thấy rằng nó không phải là loài đơn huyệt cơ bản (nguyên thủy, tổ tiên) mà là một loài thú mỏ vịt tiến bộ, và do đó, dòng dõi thú mỏ vịt và tachyglossidae tách ra sớm hơn ta tưởng. Tuy nhiên, một nghiên cứu gần đây hơn (2009) đã gợi ý rằng, trong khi Teinolophos là một loại thú mỏ vịt, nó vẫn là một thú đơn huyệt cơ bản và trước cả sự lan rộng của loài thú đơn huyệt hiện đại. Lối sống bán thủy sinh của thú mỏ vịt đã giúp chúng không bị thay thế bởi các thú có túi sau này di cư đến Úc hàng triệu năm trước, vì con của chúng có thể bị chết đuối nên con mẹ phải cho con của nó bám vào. Bằng chứng di truyền đã xác định rằng tachyglossidae chuyển hướng từ dòng dõi thú mỏ vịt 19-48 triệu năm trước, khi chúng chuyển đổi từ lối sống bán thủy sinh sang sống trên cạn.
Thú Đơn huyệt có một số đặc điểm được thừa hưởng từ tổ tiên phân bộ Một răng chó:
- Giống như thằn lằn và chim, chúng sử dụng cùng một lỗ để đi tiểu, đại tiện và sinh sản ("đơn huyệt" có nghĩa là "một lỗ").
- chúng đẻ trứng có da và không có calci, giống như của thằn lằn, rùa và cá sấu.
Không giống như các động vật có vú khác, thú Đơn huyệt không có núm vú và nuôi con bằng cách "đổ mồ hôi" sữa từ các tuyến trên bụng của chúng.
Những đặc điểm này không thể nhìn thấy trong hóa thạch và các đặc điểm chính từ quan điểm của các nhà cổ sinh vật học là:
- một xương răng mảnh khảnh trong đó vẹt hình mỏ quạ nhỏ hoặc không tồn tại.
- lỗ mở của tai nằm ở đăng sau phía đáy sọ.
- xương má nhỏ hoặc không tồn tại.
- một vòng ngực nguyên thủy với các yếu tố bụng mạnh mẽ: xương quạ, xương đòn và xương ở giữa xương ngực và xương đòn. Lưu ý: động vật có vú Theria không có xương thứ ba.
- chân nằm ngang hoặc nửa nằm ngang.
Multituberculate
Multituberculate (được đặt tên do chúng sở hữu răng hàm với nhiều loại núm) thường được gọi là "loài gặm nhấm của đại Trung Sinh", đây là một ví dụ tiêu biểu cho tiến hóa hội tụ chứ chúng thực chất không liên quan đến Bộ Gặm nhấm. Chúng tồn tại khoảng 120 triệu năm trước - lịch sử hóa thạch dài nhất của bất kỳ dòng dõi động vật có vú nào - nhưng cuối cùng bị tuyệt chủng trong thời kỳ đầu Oligocene do cạnh tranh với các loài gặm nhấm vượt trội hơn.
Một số tác giả đã thách thức cây phát sinh loài ở trên. Họ loại trừ các multituberculate khỏi nhóm chỏm cây. Theo họ thì chúng có quan hệ xa so với lớp thú ngày nay, hơn cả nhóm Morganucodontidae. Multituberculate giống với thú có vú trong đó các khớp hàm của chúng chỉ bao gồm xương răng và xương vảy - còn xương vuông và xương khớp là một phần của tai giữa; răng của chúng đa dạng, có khớp cắn và có các núm giống như động vật có vú; họ có một răng má; và cấu trúc của xương chậu gợi ý rằng chúng sinh ra các con non nhỏ bé và bất lực, giống như thú có túi hiện đại. Mặt khác, chúng khác với các động vật có vú hiện đại ở nhiều đặc điểm như:
- "Các răng hàm" của chúng có hai dãy núm song song, không giống cấu trúc răng hàm ba núm của các loài thú chỏm cây thực sự.
- Hành động nhai ở lớp thú chỏm cây là cơ chế nghiền thức ăn từ bên sang bên, có nghĩa là khớp cắn của chúng chỉ ở một bên hàm vào bất kì thời điểm nào, trong khi đó hàm multituberculate không hỗ trợ cơ chế nhai đó. Thay vào đó, khi đóng hàm lại, răng dưới của chúng được kéo về sau trượt qua răng trên giúp chúng nghiền thức ăn.
- Phần trước xương má tạo bởi phần lớn từ xương hàm trên thay vì xương gò má.
- Xương vảy không tạo nên hộp sọ
- Mõm không giống như động vật có vú chỏm cây; trên thực tế, nó trông giống như của một loài pelycosaur, chẳng hạn như Dimetrodon. Mõm multituberculate hình hộp, với mõm trên phẳng rộng tạo thành hai bên, đỉnh mũi và mảnh tiền mõm trên cao ở phía trước.
Theria
Theria (nghĩa là "thú") là một nhánh chứa tổ tiên chung cuối cùng của hai nhóm Eutheria và Metatheria. Các đạc điểm chung bao gồm:
- không sở hữu xương ở giữa xương ngực và xương đòn.
- không có xương quạ hoặc chúng được kết hợp với xương vai tạo thành mỏm vai.
- một khớp mắt cá chân với phần chính nằm giữa xương sên và xương chày, xương gót không trực tiếp kết nối với xương chày nhưng có chỗ cho các cơ bắp bám vào. (ở cá sấu thì khớp này hoạt động theo cách khác, với sự uốn được tạo ra bởi xương gót và xương sến)
- cấu trúc răng ba núm.
Metatheria
Metatheria hiện nay chỉ bao gồm các thú có túi. Một vài chi hóa thạch, như Asiatherium, có thể là một thú có túi hoặc thuộc nhánh Metatheria khác.
Một đặc điểm dễ dàng nhận diện nhất của thú có túi là hình thức sinh sản của chúng:
- Mẹ của chúng phát triển một túi noãn hoàng trong tử cung của nó để cung cấp chất dinh dưỡng đến phôi. Phôi của các bandicoot, gấu túi và gấu túi mũi trần cũng hình thành các cơ quan giống nhau thai kết nối chúng với thành tử cung, mặc dù cơ quan giống nhau thai nhỏ hơn ở thú có nhau thai và ta vẫn không chắc chắn rằng chúng cũng có vai trò chuyển chất dinh dưỡng từ mẹ sang phôi.
- Thời kì mang thai rất ngắn, thường là bốn đến năm tuần. Phôi được sinh ra ở giai đoạn phát triển rất sớm và thường chỉ dài dưới 5,1 cm sau khi sinh. Có ý kiến cho rằng việc mang thai ngắn là cần thiết để giảm nguy cơ hệ thống miễn dịch của người mẹ sẽ tấn công phôi thai.
- Con thú có túi sơ sinh sẽ sử dụng bàn chân trước (với bàn tay tương đối khỏe) để trèo lên núm vú, thường nằm trong túi trên bụng của mẹ. Người mẹ cho con nhỏ ăn bằng cách co thắt các cơ trên tuyến vú, vì con non quá yếu để bú. Nhu cầu của thú có túi sơ sinh cần sử dụng chân trước để leo lên núm vú theo lịch sử được cho là đã hạn chế sự tiến hóa của nhóm metatheria, vì các chuyên gia cho rằng chân trước của chúng không thể biến hóa thành các cấu trúc chuyên biệt như cánh, móng hoặc chân chèo. Tuy nhiên, một số laòi bandicoot, đáng chú ý nhất là bandicoot chân lợn, có móng guốc rất giống với móng guốc của thú có nhau thai, và nhiều thú lượn có túi đã phát triển.

Mặc dù một số loài thú có túi trông rất giống thú nhau thai (thylacine, "hổ có túi" hay "sói có túi" là một ví dụ điển hình), bộ xương của thú túi có một số đặc điểm phân biệt chúng với thú nhau thai:
- Một số, bao gồm thylacine, có bốn răng hàm; trong khi không có thú nhau thai nào được biết đến là có nhiều hơn ba.
- Tất cả đều có một cặp cửa sổ vòm miệng, các lỗ mở giống như cửa sổ ở dưới cùng của hộp sọ (cộng thêm các lỗ mũi nhỏ hơn).
Thú có túi cũng có một cặp xương túi (đôi khi được gọi là "xương trên mu"), hỗ trợ cái túi ở con cái. Nhưng xương này không đặc hữu đối với thú có túi, vì chúng đã được tìm thấy trong các hóa thạch của multituberculate, đơn huyệt và thậm chí cả eutheria - vì vậy chúng có thể là một đặc điểm tổ tiên phổ biến biến mất sau khi thú có túi và nhau thai tách rẽ. Một số nhà nghiên cứu nghĩ rằng chức năng ban đầu của xương trên mu là củng cố sự vận động bằng cách hỗ trợ một số cơ kéo đùi về phía trước.
Eutheria
Thời điểm xuất hiện của những loài eutheria sớm nhất là một vấn đề tranh cãi. Hóa thạch của Juramaia đã xuất hiện từ 160 triệu năm trước và được phân loại là eutheria.[63] Hóa thạch của Eomaia từ 125 triệu năm trước trong kì Phấn trắng sớm đã được phân loại là eutherian. Tuy nhiên, một phân tích gần đây về các gen tinh trạng đã phân loại Eomaia là một loài tiền eutheria và báo cáo rằng các mẫu vật eutheria sớm nhất đến từ Maelestes, có niên đại vào 91 triệu năm trước. Nghiên cứu đó cũng báo cáo rằng eutheria không đa dạng hóa đáng kể cho đến sau sự tuyệt chủng thảm khốc ở ranh giới kỉ Phân Trắng - Cổ Cận, khoảng 66 triệu năm trước.
Eomaia đã được tìm thấy có một số đặc điểm của thú có túi và các metatheria trước đó:
- Xương trên mu kéo dài về phía trước từ xương chậu, không được tìm thấy trong bất kỳ thú nhau thai hiện đại nào, nhưng được tìm thấy trong tất cả các động vật có vú khác - các mammaliaform sớm, eutheria không có nhau thai, thú có túi và thú đơn huyệt - cũng như trong các Cung thú thuộc Phân bộ Một răng chó gần gũi với thú có vú. Chức năng của chúng là làm cứng cơ thể trong quá trình vận động. Sự cứng lại này sẽ có hại cho nhau thai, mà cần được mở rộng.
- Một khe chậu hẹp, chỉ ra rằng con non rất bé khi sinh và có thai kỳ ngắn, như trong thú có túi hiện đại. Điều này cho thấy nhau thai là một sự phát triển sau này.
- Năm răng cửa ở mỗi bên hàm trên. Con số này là điển hình của metatheria, và số lượng tối đa trong các thú nhau thai là ba, ngoại trừ các homodont, chẳng hạn như Dasypodidae. Song tỷ lệ răng hàm so với răng tiền hàm của Eomaia (nó có nhiều răng hàm hơn răng hàm) là điển hình của eutheria, bao gồm cả thú nhau thai, và là con số không bình thường trong thú có túi.
Một đặc điểm dễ dàng nhận diện của thú có nhau thao là hình thức sinh sản của chúng:
- Phôi tự gắn vào tử cung thông qua nhau thai lớn qua đó mẹ cung cấp thức ăn và oxy và loại bỏ các chất thải.
- Thời kỳ mang thai tương đối dài và con non khá phát triển khi sinh. Ở một số loài, (đặc biệt là các động vật ăn cỏ sống trên đồng bằng) con non đã có thể chạy trong vòng 1 giờ sau sinh.
Sự mở rộng của thú có vú trong đại Trung Sinh
Vẫn còn nhiều cái đúng trong hình mẫu dập khuôn coi thú có vú trong Đại Trung Sinh là những "loài ăn côn trùng nhỏ sống về đêm", nhưng những phát hiện gần đây, chủ yếu ở Trung Quốc, cho thấy một số mammaliaform và động vật có vú chỏm cây lớn hơn nhiều và có các lối sống khác nhau. Ví dụ:
- Fruitafossor, từ cuối kỷ Jura khoảng 150 triệu năm trước, có kích thước bằng một con sóc chuột và răng của nó, và dường như dùng để phá vỡ tổ của côn trùng xã hội.[73]
- Tương tự, loài gobiconodontid Spinolestes có các đặc điểm thích nghi giống với các loài hóa thạch và hội tụ với các xenarthra nhau thai như vảy và đốt sống xen kẽ, do đó, nó cũng có thể có thói quen giống như thú ăn kiến. Nó cũng đáng chú ý bởi sự hiện diện của gai giống của Acomys hiện đại.
- Volaticotherium, từ ranh giới kỷ Phấn trắng khoảng 125 triệu năm trước, là loài động vật có vú bay lượn được biết đến sớm nhất và có một màng cánh kéo dài giữa các chi của nó, giống như một con sóc bay hiện đại. Điều này cũng cho thấy rằng nó hoạt động vào ban ngày.[74] Argentoconodon có liên quan chặt chẽ cũng cho thấy sự thích nghi tương tự.[75]
- Repenomamus, một loài eutrichodont từ kì Phấn trắng sớm 130 triệu năm trước, là một loài săn mồi có hình dáng giống con lửng, đôi khi ăn khủng long non. Hai loài đã được công nhận, một có chiều dài hơn 1 m và nặng khoảng 12–14 kg, loài kia dài chưa đến 0,5 m và nặng 4 – 6 kg.[76][77]
- Ichthyoconodon, một loài eutriconodont từ Berriasian của Maroc, hiện được biết đến từ các răng dạng hàm được tìm thấy trong các trầm tích biển. Những chiếc răng này có núm nhọn và giống với thú có vú ăn cá, và không giống với thú có vú chúng không bị mòn đi, vì vậy xương của nó đã có thể bị chôn tại chỗ hoặc gần đó. Đây là một thú biển, một trong những ví dụ được biết đến từ đại Trung Sinh.[81] Mặt khác, đây cũng có thể là một động vật có vú biết bay.[75]
Sự tiến hóa các đặc điểm của lớp Thú
Hàm và tai giữa
Hadrocodium, hóa thạch có niên đại từ thế Jura sớm, cung cấp bằng chứng rõ ràng đầu tiên về khớp hàm và tai giữa của động vật có vú thực sự, trong đó khớp hàm được hình thành bởi xương răng và xương squamosal trong khi xương khớp và xương vuông di chuyển lên tai giữa, nơi chúng được gọi là xương đe và xương búa.
Một phân tích về chi Teinolophos đơn huyệt cho thấy loài động vật này có khớp hàm trước thú có vú được hình thành bởi xương góc và xương vuông và tai giữa của thú có vú rõ ràng tiến hóa hai lần hoàn toàn độc lập, trong các loài đơn huyệt và ở theria, nhưng ý tưởng này bị tranh cãi. Trên thực tế, hai trong số các tác giả của đề xuất này đã đồng xuất bản một bài báo diễn giải lại các đặc điểm tương tự như bằng chứng cho thấy Teinolophos là một thú mỏ vịt đầy đủ, có nghĩa là nó sẽ có khớp hàm và tai giữa của động vật có vú.
Tiết sữa
Có ý kiến cho rằng chức năng ban đầu của tuyến sữa là giữ ẩm cho trứng. Phần lớn lập luận dựa trên các loài Đơn huyệt (động vật có vú đẻ trứng):
- Trong khi trứng ối thường được mô tả là giúp cho các sinh vật ban đầu ra khỏi nước, hầu hết trứng bò sát thực sự cần độ ẩm nếu không chúng sẽ bị khô.
- Thú Đơn huyệt không có núm vú, nhưng tiết ra sữa từ một mảng lông trên bụng của chúng.
- Trong quá trình ấp, trứng Đơn huyệt được bao phủ trong một chất dính mà nguồn gốc không được biết đến. Trước khi đẻ trứng, vỏ của chúng chỉ có ba lớp. Sau đó, một lớp thứ tư xuất hiện với thành phần khác với lớp thứ ba. Chất dính và lớp thứ tư này có thể đã được sản xuất bởi các tuyến vú.
- Nếu vậy, điều đó có thể giải thích tại sao các mảng mà từ đó các loài thú đơn huyệt tiết ra sữa có lông. Nó dễ dàng lan truyền độ ẩm và các chất khác cho trứng từ một khu vực rộng, nhiều lông, hơn là từ một núm vú nhỏ, trần trụi.
Nghiên cứu sau đó đã chứng minh rằng casein đã xuất hiện ở tổ tiên động vật có vú phổ biến vào khoảng từ 200-310 triệu năm trước. Câu hỏi về việc liệu một chất tiết ra để giữ ẩm cho trứng được chuyển thành chức năng tiết sữa thực sự vẫn bỏ ngỏ. Một thú có vú nhỏ gọi là Sinocodon, thường được coi là nhóm chị em của tất cả các động vật có vú sau này, có răng cửa ở cả những cá thể nhỏ nhất. Kết hợp với một hàm lỏng lẻo, rất có thể chúng không bú được. Do đó, con nhỏ biết bú có thể đã tiến hóa ngay tại giai đoạn chuyển đổi trước động vật có vú/động vật có vú. Tuy nhiên, tritylodontid, thường được coi là cơ bản hơn, cho thấy bằng chứng về con bú. Morganucodontan, cũng được coi là một loài thuộc nhóm Mammaliaforme cơ bản, cũng có bằng chứng cho thấy khả năng tiết sữa.
Tóc và lông mao
Bằng chứng rõ ràng đầu tiên về tóc hoặc lông mao là trong hóa thạch của Castorocauda và Megaconus, từ 164 triệu năm trước ở giữa kỷ Jura.[37] Vì cả hai loài động vật có vú Megaconus và Castorocauda đều có bộ lông kép, với cả lông bảo vệ và lớp lông lót, có thể giả định rằng tổ tiên chung cuối cùng của chúng cũng sơ hữu đặc điểm như vậy. Loài vật này hẳn là từ kỷ Tam Điệp vì nó là tổ tiên của Tikitherium. Gần đây, việc phát hiện ra tàn dư của tóc trong phân hóa thạch từ kỷ Permi đẩy lùi nguồn gốc của lông động vật có vú sớm hơn nhiều trong dòng động vật Một cung bên thành Bộ Cung thú ở đại Cổ Sinh.
Vào giữa những năm 1950, một số nhà khoa học đã giải thích lỗ thông ở hàm trên và mảnh tiền hàm trên của cynodont là các kênh cung cấp mạch máu và dây thần kinh cho bộ cảm nhận rung động (râu ria) và cho rằng đây là bằng chứng về tóc hoặc lông. Tuy nhiên, người ta đã sớm chỉ ra rằng các lỗ thông không nhất thiết phải cho thấy rằng một con vật có cơ quan cảm thụ rung động; loài thằn lằn hiện đại Tupinambis có lỗ thông gần giống với Thrinaxodon.
Tuy nhiên, các nguồn phổ biến vẫn tiếp tục gán râu cho Thrinaxodon. Một hóa thạch dấu vết từ thế Hạ Tam Điệp đã bị coi nhầm là dấu chân của cynodont cho thấy tóc, nhưng cách giải thích này đã bị bác bỏ. Một nghiên cứu về các khe hở sọ cho thấy các dây thần kinh mặt kết nối với râu ở các động vật có vú còn tồn tại cho thấy Prozostrodontia, tổ tiên nhỏ của động vật có vú, cho thấy râu ria tương tự như động vật có vú, nhưng các Cung thú kém tiên tiến hơn sẽ không có râu hoặc râu không năng động. Lông có thể đã tiến hóa từ râu ria. Bản thân râu ria có thể đã tiến hóa như một phản ứng với lối sống về đêm và/hoặc đào hang.
Ruben & Jones (2000) lưu ý rằng các tuyến Harderia, tiết ra lipid để bọc lông, đã có mặt ở các thú có vú sớm nhất như Morganucodon, nhưng không có trong các Cung thú gần động vật có vú như Thrinaxodon. Mã gen di truyền Msx2 dùng để duy trì lông mao cũng có liên quan đến sự đóng mắt đỉnh ở động vật có vú, cho thấy lông và thiếu mắt đỉnh có liên quan. Mắt đỉnh có mặt ở Thrinaxodon, nhưng không có trong các cynognath tiên tiến hơn (Probainognathia).
Máu nóng
"Máu nóng" là một thuật ngữ mơ hồ, do nó chỉ một trong số các kiểu hệ trao đổi chất dưới đây:
- Nội nhiệt, khả năng tạo nhiệt bên trong hơn là lấy nhiệt thông qua các hành vi như phơi nắng hay từ hoạt động cơ bắp.
- Đẳng nhiệt, duy trì nhiệt độ cơ thể khá ổn định. Hầu hết các enzyme có nhiệt độ hoạt động tối ưu; hiệu quả giảm nhanh chóng ngoài phạm vi chuyên biệt. Một sinh vật đẳng nhiệt nhỏ chỉ cần một vài các enzyme hoạt động trong vùng nhiệt độ nhỏ.
- Trao đổi chất nhanh. duy trì tốc độ trao đổi chất nhanh, nhất là lúc nghỉ. Điều này đòi hỏi nhiệt độ cơ thể khá cao và ổn định do hiệu ứng Q10: các quá trình sinh hóa bị chậm đi khoảng một nửa nếu thân nhiệt của động vật giảm 10 °C.
Vì các nhà khoa học không thể biết về cơ chế bên trong của các sinh vật đã tuyệt chủng, hầu hết các cuộc thảo luận tập trung vào hệ trao đổi chất nhanh và đẳng nhiệt. Tuy vậy, người ta thường đồng ý rằng hệ nội nhiệt tiến hóa lần đầu ở các loài không phải động vật có vú như dicynodont, có tỷ lệ cơ thể liên quan đến sự giữ nhiệt, xương mạch máu với kênh Haversia, và có thể đa mọc lông mao. Gần đây, có ý kiến cho rằng trao đổi chất nội nhiệt đã phát triển từ xa xưa ở chi Ophiacodon.
Tham khảo
Liên kết ngoài
- The Cynodontia Lưu trữ 2008-10-12 tại Wayback Machine bàn về nhiều khía cạnh tiến hóa từ các cynodont tới động vật có vú, với rất nhiều dẫn nguồn tham khảo.
- Mammals, đàm thoại với Richard Corfield, Steve Jones & Jane Francis trên BBC Radio 4 (In Our Time, ngày 13 tháng 10, 2005)