
| Virus | |
|---|---|

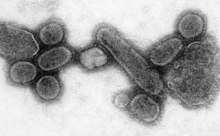
SARS-CoV-2, một nhánh của phân họ Coronavirinae
| |
| Phân loại virus | |
| Nhóm: I–VII | |
| Nhóm | |
I: Virus dsDNA |
Virus, thường được viết là vi-rút (bắt nguồn từ tiếng Pháp virus /viʁys/), còn được gọi là siêu vi, siêu vi khuẩn hay siêu vi trùng, là một tác nhân truyền nhiễm chỉ nhân lên được khi ở bên trong tế bào sống của một sinh vật khác. Virus có thể xâm nhiễm vào tất cả các dạng sinh vật, từ động vật, thực vật cho tới vi khuẩn và cổ khuẩn. Kể từ bài viết đầu tiên của Dmitriy Iosifovich Ivanovsky năm 1892, mô tả về một dạng mầm bệnh không thuộc vi khuẩn mà lây nhiễm vào cây thuốc lá, và sự khám phá ra virus khảm thuốc lá của Martinus Beijerinck năm 1898, cho đến nay có khoảng 9,000 loại virus đã được miêu tả chi tiết, mặc dù vẫn còn có tới hàng triệu (hoặc tỷ) dạng virus khác nhau. Virus được tìm thấy ở hầu hết mọi hệ sinh thái trên Trái Đất và là dạng có số lượng nhiều nhất trong tất cả các thực thể sinh học. Khoa học nghiên cứu virus được biết với tên virus học (virology), một chuyên ngành phụ của vi sinh.
Các phần tử (hay hạt) virus (được gọi là virion hoặc vi-ri-ông) được tạo thành từ hai hoặc ba bộ phận: phần vật chất di truyền được tạo nên từ DNA hoặc RNA – những phân tử dài có mang thông tin di truyền, một lớp vỏ protein – được gọi với tên capsid – có chức năng bảo vệ hệ gen và một lớp vỏ bọc bên ngoài làm từ lipid mà bao bọc bên ngoài lớp vỏ protein khi virus ở ngoài tế bào (chỉ có trong một số trường hợp). Hình dạng của virus có sự khác nhau, từ dạng xoắn ốc hay khối hai mươi mặt đều đơn giản cho tới những cấu trúc phức tạp hơn. Một virus có kích thước trung bình vào khoảng 1/100 kích cỡ trung bình của một con vi khuẩn. Hầu hết virus đều quá nhỏ nên không thể quan sát trực tiếp dưới kính hiển vi quang học.
Nguồn gốc của virus trong lịch sử tiến hóa của sự sống không rõ ràng: một số có thể đã tiến hóa từ những plasmid – những đoạn DNA ngắn có khả năng di chuyển giữa các tế bào – trong khi số khác có thể đã tiến hóa từ vi khuẩn. Trong tiến hóa, virus là một phương tiện chuyển gen ngang quan trọng, góp phần gia tăng sự đa dạng di truyền. Virus được công nhận là một dạng sống bởi một số nhà khoa học, do chúng có mang vật chất di truyền, có thể sinh sản và tiến hóa thông qua quá trình chọn lọc tự nhiên. Tuy nhiên mặt khác chúng lại thiếu những đặc tính thiết yếu (như cấu trúc tế bào) – những điều được công nhận rộng rãi là cần thiết để được coi như sinh vật sống. Bởi vì chỉ có một số chứ không tất cả các phẩm chất cần thiết, nên virus được mô tả như "những sinh vật ở bên lề của sự sống". Tuy nhiên, virus chỉ có thể xâm nhập qua một số tế bào nhất định nhờ có giác bám (gai glycoprotein) của virus bám đặc hiệu lên thụ thể của tế bào chủ.
Virus lây lan theo nhiều cách; virus thực vật thường được truyền từ cây này sang cây khác qua những loài côn trùng hút nhựa cây như rệp vừng; trong khi virus động vật lại có thể được truyền đi nhờ những côn trùng hút máu. Những sinh vật mang mầm bệnh như vậy được gọi là những véc-tơ. Virus cúm lan truyền thông qua ho và hắt hơi. Norovirus và rotavirus, nguyên nhân chính của bệnh viêm dạ dày ruột siêu vi, lây lan qua đường phân-miệng và truyền từ người này sang người khác thông qua tiếp xúc, cũng như xâm nhập vào cơ thể qua thức ăn hay nước uống. HIV là một trong vài loại virus lây nhiễm thông qua quan hệ tình dục và tiếp xúc với máu bị nhiễm bệnh. Mỗi virus chỉ có thể xâm nhiễm vào một số dạng tế bào vật chủ nhất định, gọi là "biên độ vật chủ" (host range); biên độ này có thể rất hẹp hoặc rất rộng, tùy vào số lượng những sinh vật khác nhau mà virus có khả năng lây nhiễm.
Sự xâm nhập của virus trong động vật đã kích hoạt một phản ứng miễn dịch nhằm loại bỏ virus xâm nhiễm. Những phản ứng miễn dịch cũng có thể được tạo ra bởi vắc-xin, giúp tạo ra miễn dịch chủ động nhân tạo đối với một virus xâm nhiễm nhất định. Tuy nhiên, một số virus, bao gồm những loại gây ra AIDS và viêm gan siêu vi, lại có thể trốn tránh những phản ứng trên và gây ra sự nhiễm bệnh mãn tính. Đa phần các chất kháng sinh không có hiệu quả đối với virus, dù vậy cũng đã có những loại thuốc kháng virus được phát triển.
Từ nguyên
Từ virus trong tiếng Việt bắt nguồn từ từ tiếng Pháp virus /viʁys/.. Từ virus trong tiếng Pháp thì bắt nguồn từ từ tiếng Latinh virus, có nghĩa là chất độc.Virus được ghi nhận với nghĩa "tác nhân gây bệnh truyền nhiễm" lần đầu vào năm 1728, trước khi Dmitri Ivanovsky phát hiện ra virus vào năm 1892. Thuật ngữ virion bắt đầu có từ năm 1959, được dùng để chỉ một phần tử virus, đơn lẻ, ổn định, có khả năng lây nhiễm được giải phóng ra từ tế bào và hoàn toàn có thể xâm nhập các tế bào khác cùng loại.
Lịch sử

Vào những năm 1880, Louis Pasteur khi nghiên cứu về bệnh dại đã không thể tìm ra tác nhân gây ra bệnh này, và dự đoán về một mầm bệnh quá nhỏ để có thể phát hiện được dưới kính hiển vi. Năm 1884, nhà vi sinh vật học người Pháp Charles Chamberland đã phát minh ra một bộ lọc (được biết tới ngày nay là bộ lọc Chamberland hay bộ lọc Chamberland-Pasteur) với các lỗ có kích thước còn nhỏ hơn cả vi khuẩn. Nhờ thế, ông có thể cho một dung dịch chứa vi khuẩn chảy qua bộ lọc và hoàn toàn loại bỏ chúng khỏi dung dịch. Năm 1892, nhà sinh vật học người Nga, Dmitriy Iosifovich Ivanovskiy đã sử dụng bộ lọc này để nghiên cứu về thứ mà hiện nay được biết với tên virus khảm thuốc lá. Thí nghiệm của ông cho thấy chiết xuất từ lá cây thuốc lá nhiễm bệnh được nghiền nát vẫn có thể lây nhiễm sau khi lọc. D. I. Ivanovskiy đề xuất rằng sự nhiễm bệnh có thể là do một độc tố từ vi khuẩn gây ra, nhưng đã không theo đuổi ý tưởng đó. Lý do là vì vào thời điểm đó, người ta đã nghĩ rằng tất cả các tác nhân truyền nhiễm đều có thể bị các bộ lọc giữ lại và chỉ có thể phát triển trên một môi trường dinh dưỡng – đây là một phần của thuyết mầm bệnh. Năm 1898, nhà vi sinh vật học người Hà Lan Martinus Beijerinck đã lặp lại thí nghiệm và tin rằng dung dịch đã lọc vẫn còn chứa một dạng tác nhân truyền nhiễm mới. Ông nhận thấy rằng tác nhân này chỉ có thể nhân lên trong tế bào đang phân chia, nhưng thí nghiệm của ông không chỉ ra là nó được làm từ các hạt; ông gọi tác nhân này là một contagium vivum fluidum (mầm sống có thể hòa tan) và sử dụng lại từ virus để gọi nó. Beijerinck giữ quan điểm rằng virus có bản chất là chất lỏng, tuy nhiên về sau thuyết này đã bị bác bỏ bởi Wendell Stanley, người chứng minh được chúng có dạng hạt. Trong cùng năm đó, Friedrich Loeffler và Frosch đã cho chảy lần đầu tiên dung dịch chứa virus động vật – tác nhân gây bệnh lở mồm long móng (aphthovirus) – qua một bộ lọc tương tự.
Vào đầu của thế kỷ XX, Frederick Twort - một nhà vi khuẩn học người Anh - đã khám phá ra một nhóm những virus có thể xâm nhiễm vào vi khuẩn, mà nay gọi là thực khuẩn thể (bacteriophage hay phage), và nhà vi sinh học người Canada gốc Pháp Félix d'Herelle đã miêu tả về virus rằng: khi thêm chúng vào vi khuẩn trên thạch agar, sẽ tạo ra những vùng vi khuẩn bị chết. Ông đã pha loãng chính xác một dịch huyền phù những virus trên và khám phá ra rằng những dịch pha loãng cao nhất (mật độ virus thấp nhất), thay vì giết chết toàn bộ vi khuẩn, đã tạo những vùng riêng biệt gồm những cá thể bị chết. Tính toán diện tích những vùng này và nhân với hệ số pha loãng cho phép ông tính được số lượng virus trong dịch huyền phù gốc. Phage đã được cho rằng sẽ là một giải pháp điều trị tiềm năng cho những bệnh như thương hàn và tả, nhưng triển vọng của chúng đã bị lãng quên cùng với sự phát triển của penicillin. Nghiên cứu về phage đã cung cấp cái nhìn sâu hơn về sự bất hoạt và kích hoạt gen, và một cơ chế hữu hiệu cho việc đưa những gen bên ngoài vào bên trong vi khuẩn.
Cho đến cuối thế kỷ XIX, virus được định nghĩa dựa trên sự lây nhiễm của chúng, khả năng chống lọc, và việc chúng đòi hỏi phải có một vật chủ. Virus đã từng chỉ được nuôi trong thực vật và động vật. Năm 1906, Ross Granville Harrison phát minh ra một phương pháp để nuôi dưỡng mô trong bạch huyết, và sau đó năm 1913, E. Steinhardt, C. Israeli, và R. A. Lambert đã sử dụng phương pháp này để phát triển virus vaccinia trong những mảnh vụn của mô giác mạc chuột lang nhà. Năm 1928, H. B. Maitland và M. C. Maitland đã nuôi virus vaccinia trong những thể huyền phù của thận gà băm nhỏ. Phương pháp của họ đã không được áp dụng rộng rãi cho tới thập niên 1950, khi poliovirus được nuôi ở quy mô lớn phục vụ việc sản xuất vaccine.
Một bước đột phá khác đến vào năm 1931, khi nhà bệnh học Hoa Kỳ Ernest William Goodpasture đã nuôi dưỡng virus cúm và vài loại virus khác trong trứng gà đã thụ tinh. Năm 1949, John Franklin Enders, Thomas Weller, và Frederick Robbins cũng nuôi cấy virus bại liệt trong tế bào phôi người, virus đầu tiên được nuôi mà không sử dụng mô thể rắn của động vật hay trứng. Công trình này cho phép Jonas Salk tạo ra một vắc-xin bại liệt hiệu quả.
Những hình ảnh đầu tiên của virus thu nhận được là nhờ sự phát minh ra kính hiển vi điện tử năm 1931 của hai kĩ sư người Đức Ernst Ruska và Max Knoll. Năm 1935, Wendell Meredith Stanley - một nhà sinh hóa và virus học người Mỹ - đã nghiên cứu virus khảm thuốc lá và nhận thấy chúng được tạo thành phần lớn từ protein. Một thời gian ngắn sau, virus này đã được phân tách thành các phần protein và RNA riêng biệt. Virus khảm thuốc lá là dạng virus đầu tiên được tinh thể hóa và cấu trúc của nó do đó đã được làm sáng tỏ chi tiết. Những hình ảnh nhiễu xạ tia X đầu tiên của virus kết tinh đã được Bernal và Fankuchen thu được vào 1941. Dựa trên những tấm hình này, Rosalind Franklin đã khám phá ra cấu trúc DNA hoàn thiện của loại virus này vào năm 1955. Cùng trong năm đó, Heinz Fraenkel-Conrat và Robley Williams chứng minh được là chiết xuất RNA và vỏ protein của virus khảm thuốc lá có thể tự lắp ráp lại để tạo thành những virus có chức năng, cho thấy cơ chế đơn giản này có thể là cách virus sinh ra trong tế bào vật chủ.
Nửa sau của thế kỷ XX đánh dấu một kỉ nguyên vàng cho sự khám phá virus với hầu hết trong số hơn 2.000 loài virus động vật, thực vật và vi khuẩn đã được phát hiện trong những năm này. Năm 1957, virus arteri ở ngựa và virus gây bệnh tiêu chảy ở bò (một loại pestivirus) đã được phát hiện. Năm 1963, virus viêm gan siêu vi B cũng được Baruch Blumberg khám phá, và năm 1965, Howard Temin đã mô tả loại retrovirus đầu tiên. Sau đó, enzym phiên mã ngược (Reverse transcriptase), loại enzym quan trọng mà retrovirus sử dụng để phiên mã RNA của chúng thành DNA, được miêu tả lần đầu vào năm 1970, một cách độc lập bởi Howard Martin Temin và David Baltimore. Năm 1983, nhóm nghiên cứu của Luc Montagnier tại Viện Pasteur ở Pháp, đã lần đầu tiên phân lập một loại retrovirus được biết với tên gọi ngày nay là virus suy giảm miễn dịch ở người (HIV).
Nguồn gốc
Virus được tìm thấy ở bất cứ nơi nào có sự sống và có thể đã tồn tại kể từ khi tế bào sống đầu tiên được tiến hóa thành. Nguồn gốc của virus không rõ ràng bởi chúng không tạo hóa thạch, vì vậy các kĩ thuật phân tử đã được sử dụng để so sánh RNA hay DNA của virus và là một công cụ hiệu quả để nghiên cứu xem chúng phát sinh ra sao. Nhìn chung, có ba giả thuyết chính để giải thích nguồn gốc của virus:
Giả thuyết thoái hóa
Giả thuyết này cho rằng virus có thể đã từng là những tế bào nhỏ ký sinh bên trong những tế bào lớn hơn. Trải qua thời gian, những gen không cần thiết cho sự ký sinh này mất đi. Những vi khuẩn như Rickettsia và Chlamydia cũng sống trong tế bào và giống như virus, chỉ có thể sinh sản khi ở bên trong tế bào vật chủ. Những vi khuẩn này đã hỗ trợ cho giả thuyết thoái lui, do sự phụ thuộc ký sinh của chúng có thể đã làm mất đi những gen cho phép chúng tồn tại bên ngoài tế bào. Giả thuyết này còn được gọi là "giả thuyết thoái hóa" (degenracy hypothesis), hoặc "giả thuyết suy giảm" (reduction hypothesis).
Giả thuyết nguồn gốc từ tế bào
Theo giả thuyết này, một số virus có thể đã tiến hóa từ những mảnh DNA hay RNA mà "thoát ra" (escape) từ hệ gen của những sinh vật lớn hơn. DNA thoát ra có thể là từ những plasmid (những đoạn DNA trần mà có thể di chuyển giữa những tế bào) hoặc từ những transposon (những phân tử DNA mà nhân lên và di chuyển quanh những vị trí khác nhau bên trong bộ gen của tế bào). Từng được gọi là những "gen nhảy", transposon là những ví dụ của các yếu tố di truyền di động và có thể là nguồn gốc của một số virus. Chúng được Barbara McClintock phát hiện ở cây ngô vào năm 1950. Đôi khi giả thuyết này còn được gọi là "giả thuyết lang thang" (vagrancy hypothesis), hoặc "giả thuyết trốn thoát" (escape hypothesis).
Giả thuyết đồng tiến hóa
Giả thuyết này còn được gọi là "giả thuyết virus-đầu tiên" (virus-first hypothesis), đề xuất rằng virus có thể đã tiến hóa từ những phân tử protein và acid nucleic phức tạp cùng một thời điểm khi tế bào xuất hiện lần đầu trên Trái Đất, và đã không hề bị phụ thuộc vào tế bào trong hàng tỷ năm. Viroid là những phân tử RNA mà không được phân loại là virus bởi chúng thiếu đi lớp vỏ protein. Tuy nhiên, chúng có những đặc tính tương đồng với vài loại virus và thường được gọi là những "tác nhân dưới virus" (subviral agent). Viroid là những mầm bệnh quan trọng ở thực vật. Chúng không mã hóa protein nhưng lại tương tác với tế bào chủ và sử dụng bộ máy tế bào vật chủ cho sự nhân lên của chúng. Virus viêm gan siêu vi D trên người cũng có bộ gen RNA tương đồng với viroid nhưng có lớp vỏ protein xuất xứ từ virus viêm gan B và không có khả năng tạo ra lớp vỏ của riêng chúng. Vì vậy, chúng là một loại virus khiếm khuyết và không thể tự nhân lên mà không có virus viêm gan B giúp đỡ. Tương tự như vậy, sputnik virophage cũng bị lệ thuộc vào mimivirus, loại virus lây nhiễm trên Acanthamoeba castellanii. Những virus phụ thuộc vào sự hiện diện của những loài virus khác trong tế bào vật chủ được gọi là những vệ tinh và có thể là đại diện cho một bước trung gian trong quá trình tiến hóa giữa viroid và virus.
Sự tranh cãi giữa các giả thuyết về nguồn gốc của virus
Trong quá khứ, tất cả những giả thuyết trên đều gặp phải vấn đề: giả thuyết thoái lui không giải thích được tại sao kể cả những ký sinh nội bào nhỏ nhất cũng không giống với virus ở bất kỳ góc độ nào. Giả thuyết trốn thoát không lý giải được về lớp vỏ capsid phức hợp và các cấu trúc khác của phần tử virus. Giả thuyết virus-đầu tiên thì trái với định nghĩa của virus là chúng đòi hỏi phải có tế bào chủ. Cho đến hiện nay, virus được công nhận là rất cổ xưa và có nguồn gốc mà bắt đầu từ trước cả sự rẽ nhánh của sự sống vào ba vực. Phát hiện này đã khiến những nhà virus học hiện đại phải xem xét và đánh giá lại cả ba giả thuyết cổ điển trên.
Bằng chứng về một thế giới của những tế bào RNA tổ tiên và những phân tích máy tính về trình tự DNA của virus và vật chủ đã đem lại sự hiểu biết tốt hơn về mối quan hệ tiến hóa giữa những loại virus khác nhau, và có thể giúp xác định tổ tiên của những virus hiện đại. Cho đến nay, những phân tích này vẫn chưa chứng tỏ được giả thuyết nào ở trên là đúng. Tuy nhiên, có vẻ như ít có khả năng rằng tất cả các virus hiện đang được biết có cùng một tổ tiên chung, và virus có lẽ đã phát sinh nhiều lần trong quá khứ bởi một hay nhiều cơ chế.
Prion là những phân tử protein có khả năng lây nhiễm mà không phải DNA hay RNA. Chúng có thể gây các bệnh như bệnh ngứa điên ở cừu (scrapie), bệnh viêm não thể bọt biển ở bò (bệnh bò điên) và bệnh suy mòn mãn tính ở hươu; ở con người, những bệnh gây ra bởi prion bao gồm bệnh Kuru, bệnh Creutzfeldt–Jakob, và hội chứng Gerstmann–Sträussler–Scheinker. Prion có khả năng nhân lên do một số protein có thể tồn tại ở hai hình dạng khác nhau, và prion thay đổi hình dạng thông thường của protein vật chủ thành dạng prion. Điều này sẽ phát động một phản ứng chuỗi khi mỗi protein prion biến đổi nhiều protein vật chủ thành nhiều prion hơn, và những prion mới này lại tiếp tục biến đổi càng lúc càng nhiều protein thành prion hơn nữa; và tất cả những bệnh liên quan tới prion được biết đến đều dẫn đến tử vong. Mặc dù những prion về cơ bản khá khác biệt với virus và viroid, thì sự khám phá ra chúng đã làm gia tăng niềm tin vào học thuyết rằng virus có thể đã tiến hóa từ những phân tử tự sao chép.
Sinh học
Đặc điểm cơ bản
- Virus rất nhỏ bé, kích thước mỗi cá thể chỉ trong khoảng 20 nm đến 200 nm (nhỏ hơn vi khuẩn hàng ngàn lần).
- Không có cấu tạo tế bào, không có màng kép lipid bao bọc.
- Có đời sống kí sinh bắt buộc.
- Vật chất di truyền là một trong hai loại: DNA hoặc RNA mà không có cả hai.
- Không có hệ giải mã và dịch mã.
- Không tăng kích thước (không lớn).
- Không tự di chuyển.
- Không có khả năng tự phát triển và phân chia
- Bị bất hoạt hoàn toàn khi ở ngoài vật chủ
Đặc điểm đời sống
Có những ý kiến khác nhau về việc liệu virus có phải là một dạng sống không, hay chỉ là những cấu trúc hữu cơ tương tác với những sinh vật sống. Chúng đã được mô tả là "những sinh vật bên lề của sự sống" ("organisms at the edge of life"), do chúng giống với những cơ thể sống ở chỗ chúng mang những gen và tiến hóa nhờ quá trình chọn lọc tự nhiên, và có thể sinh sản băng cách tạo ra rất nhiều bản sao của chính chúng bằng cách tự lắp ráp. Dù vậy, chúng lại không có cấu trúc tế bào, thứ mà thường được coi như đơn vị cơ bản của sự sống. Virus cũng không có hệ thống trao đổi chất của riêng chúng, và đòi hỏi phải có một tế bào chủ để tạo ra cá thể mới. Chúng do đó không thể sinh sản tự nhiên bên ngoài tế bào vật chủ – dù một số loài vi khuẩn như Rickettsia và Chlamydia được công nhận là sinh vật sống cũng có giới hạn tương tự. Những dạng sống được chấp nhận thường phải sử dụng phân chia tế bào để sinh sản, trong khi virus lại tự lắp ráp bên trong tế bào. Chúng cũng khác với sự tăng trưởng tự động của những tinh thể, do chúng được thừa hưởng những đột biến di truyền và phải chịu sự chọn lọc tự nhiên. Sự tự lắp ráp của virus trong tế bào chủ có ý nghĩa quan trọng cho việc nghiên cứu nguồn gốc sự sống, và củng cố niềm tin cho giả thuyết về việc sự sống có thể đã bắt đầu từ những phân tử hữu cơ tự lắp ráp.
Cấu trúc

Virus rất đa dạng về kích thước và hình dạng, được gọi chung là những hình thái của virus. Nhìn chung, virus có kích cỡ nhỏ hơn vi khuẩn. Hầu hết các virus được nghiên cứu có đường kính trong khoảng từ 20 đến 300 nanomet. Một số filovirus có tổng chiều dài lên tới 1400 nm; dù đường kính của chúng chỉ vào khoảng 80 nm. Đa phần các virus đều không thể quan sát được bằng kính hiển vi quang học, cho nên kính hiển vi điện tử quét và truyền qua được sử dụng để trực quan hóa những virion. Để gia tăng sự tương phản giữa virus và xung quanh, người ta phải sử dụng những "vết nhuộm" ("stain") với mật độ electron dày đặc. Những dung dịch muối của những kim loại nặng, như wolfram, có thể phân tán electron khỏi những vùng được các vết nhuộm che phủ. Nếu virion được bao bọc bằng các vết nhuộm (nhuộm dương tính), những chi tiết tỉ mỉ sẽ bị che mờ. Phương pháp nhuộm âm tính vì thế sẽ giúp khắc phục vấn đề này khi chỉ nhuộm màu môi trường nền.
Một phần tử (hạt) virus hoàn chỉnh - được gọi virion - bao gồm acid nucleic được bao bọc bởi một lớp vỏ protein bảo vệ gọi là capsid. Lớp vỏ này được tạo thành từ những tiểu đơn vị protein giống hệt nhau gọi là capsomer. Ngoài ra, virus có thể có một vỏ bọc bằng lipid có xuất xứ từ màng tế bào vật chủ. Vỏ capsid được tạo ra từ protein được dịch mã bởi bộ gen của virus và hình dạng của chúng chính là cơ sở để phân biệt về mặt hình thái học. Các tiểu đơn vị protein do virus mã hóa sẽ tự lắp ráp để tạo nên vỏ capsid, nhìn chung sẽ đòi hỏi sự có mặt của bộ gen virus. Những virus phức tạp còn mã hóa những protein trợ giúp cho quá trình xây dựng capsid của chúng. Những protein mà kết hợp với acid nucleic được biết với tên nucleoprotein, và sự kết hợp của những protein ở capsid với acid nucleic của virus được gọi là một nucleocapsid. Vỏ capsid và toàn bộ cấu trúc virus có thể được thăm dò vật lý (cơ học) thông qua kính hiển vi lực nguyên tử. Nhìn chung, có bốn hình thái virus chính:


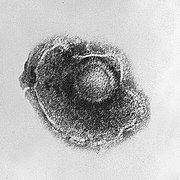
Xoắn ốc
Những virus này được tạo thành từ một loại capsomer duy nhất xếp chồng lên quanh một trục trung tâm để tạo nên cấu trúc xoắn ốc, có thể có một khoang trung tâm hoặc là một ống rỗng. Sự sắp xếp này dẫn đến việc tạo ra những virion dạng hình que hay sợi, chúng có thể ngắn và rất cứng, hoặc dài và rất linh hoạt. Vật liệu di truyền nhìn chung là RNA sợi đơn (ssRNA), nhưng đôi khi là DNA sợi đơn (ssDNA), và được gắn chặt với chuỗi xoắn protein bằng những tương tác giữa acid nucleic mang điện tích âm và phần điện tích dương trên protein. Nói chung, độ dài của vỏ capsid xoắn ốc có liên quan tới độ dài của acid nucleic bên trong nó và đường kính thì phụ thuộc vào kích thước và cách sắp xếp các capsomer. Loại virus khảm thuốc lá - mà đã được nghiên cứu kĩ lưỡng - là một ví dụ của virus dạng xoắn ốc.
Khối hai mươi mặt đều
Hầu hết virus động vật đều có dạng khối hai mươi mặt đều hoặc gần hình cầu với hai mươi mặt đều đối xứng (nhị thập diện đều). Một khối hai mươi mặt đều bình thường là cách tối ưu để tạo nên một vỏ khép kín từ những tiểu đơn vị giống y như nhau. Số lượng các capsomer tối thiểu cần đến là 12, trong đó mỗi capsomer tạo thành từ năm tiểu đơn vị y hệt nhau. Nhiều virus, ví dụ rotavirus, có nhiều hơn 12 capsomer và xuất hiện dưới dạng hình cầu nhưng vẫn giữ tính đối xứng. Capsomer tại mỗi đỉnh được bao quanh bởi 5 capsomer khác gọi là penton. Capsomer trên những mặt hình tam giác thì được bao quanh bởi 6 và gọi là hexon. Hexon về bản chất thường phẳng và penton, cấu trúc tạo nên 12 đỉnh, lại thường cong. Cùng một protein cũng có thể là tiểu đơn vị của cả penton và hexon, hoặc chúng có thể được cấu tạo bởi những protein khác nhau.
Kéo dài
Có cấu trúc một khối hai mươi mặt đều được kéo dài gấp năm lần theo chiều dài của trục; đây cũng cách sắp xếp phổ biến ở đầu của mỗi bacteriophage. Nó tạo thành một hình trụ với nắp đậy ở hai đầu.
Phức tạp
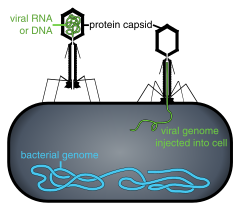
Những virus này có một capsid mà không hoàn toàn xoắn hay hoàn toàn khối hai mươi mặt đều, và có thể mang những cấu trúc thêm vào như đuôi protein hoặc một vách ngăn ngoài phức hợp. Một số bacteriophages, như Enterobacteria phage T4, có cấu trúc phức tạp bao gồm một đầu hình khối hai mươi mặt đều gắn với một đuôi xoắn; đuôi này có thể có một đĩa nền lục giác đều với các sợi đuôi protein nhô ra. Cấu trúc đuôi này đóng vai trò một ống tiêm phân tử, giúp gắn vào vi khuẩn vật chủ rồi sau đó bơm bộ gen của virus vào bên trong tế bào.
Vỏ bọc virus
Một vài loài virus bao phủ bản thân chúng bằng một lớp vỏ bọc, là một dạng chỉnh sửa của một trong những màng tế bào: có thể là lớp màng ngoài bao quanh tế bào vật chủ bị nhiễm, hoặc màng trong như màng nhân hay mạng lưới nội chất, tạo ra một lớp lipid kép bao ngoài được gọi là vỏ bọc virus (viral envelope). Màng này được đính vào những protein mã hóa bởi bộ gen virus và bộ gen vật chủ; ngoài ra, tất cả màng lipid và bất cứ carbohydrat đều có nguồn gốc từ vật chủ. Virus cúm và HIV là ví dụ cho những loại sử dụng chiến thuật này. Hầu hết các virus có màng đều phụ thuộc vào lớp vỏ của chúng để có thể xâm nhiễm vào vật chủ.
Virus lớn
poxvirus là những virus lớn, phức tạp có hình thái bất thường. Bộ gen của chúng được gắn với protein bên trong một cấu trúc đĩa ở trung tâm gọi là một nucleoid. Nucleoid được bao quanh bởi một lớp màng và hai cơ quan ở hai bên mà không rõ chức năng. Virus có một vỏ bọc ngoài với một lớp protein dày đính trên bề mặt vỏ bọc. Về tổng thể, virion của poxvirus là đa hình thái, do có thể có hình dạng trứng hoặc hình dạng gạch. Mimivirus là virus lớn nhất thường được biết, với đường kính vỏ capsid là 400 nm. Các sợi protein có thể nhô lên khỏi bề mặt với chiều dài lên tới 100 nm. Vỏ capsid có dạng đa giác đều dưới kính hiển vi điện tử, do đó có thể đây là một virus dạng khối hai mươi mặt đều. Năm 2011, các nhà nghiên cứu đã khám phá ra một virus lớn hơn ở đáy đại dương gần bờ biển Las Cruces, Chile. Nó tạm được đặt tên là Megavirus chilensis, và có thể nhìn thấy được dưới kính hiển vi quang học cơ bản.
Một số loại virus khác
Một vài loại virus mà lây nhiễm vào vi khuẩn cổ có cấu trúc phức tạp mà không liên quan tới bất kỳ dạng virus nào khác, với những hình dạng bất thường rất đa dạng, từ những cấu trúc hình con suốt, cho tới hình que có móc, hình giọt nước, hay thậm chí cả hình cái chai. Những virus trên vi khuẩn cổ giống khác thì tương tự với những bacteriophage có đuôi, và có thể có nhiều dạng cấu trúc đuôi khác nhau.
Bộ gen
| Thuộc tính | Thông số |
|---|---|
| Acid nucleic |
|
| Hình dạng |
|
| Số sợi |
|
| Tính |
|
Có rất nhiều cấu trúc bộ gen có thể được nhìn thấy ở các loài virus; chúng là nhóm có sự đa dạng cấu trúc bộ gen nhiều hơn thực vật, động vật, vi khuẩn hay vi khuẩn cổ. Có tới hàng triệu loại virus khác nhau, dù mới chỉ có khoảng 7.000 trong số đó đã được mô tả chi tiết. Cho đến tháng 1 năm 2021, cơ sở dữ liệu bộ gen virus của Trung tâm Thông tin Công nghệ sinh học Quốc gia Hoa Kỳ (NCBI) lưu trữ hơn 193.000 trình tự bộ gen hoàn chỉnh, nhưng chắc chắn còn nhiều vấn đề cần khám phá.
Một virus có các gen DNA hay RNA được gọi tương ứng là một virus DNA hoặc virus RNA. Phần lớn virus có bộ gen là RNA. Virus thực vật có xu hướng có bộ gen RNA sợi đơn và bacteriophage thường có bộ gen DNA sợi đôi.
Bộ gen virus có dạng mạch vòng, như ở polyomavirus, hay mạch thẳng, như ở adenovirus. Loại acid nucleic không liên quan tới hình dạng của bộ gen. Ở các virus RNA hay một số virus DNA, bộ gen thường được phân làm các phần riêng biệt, gọi là bị phân đoạn. Với virus RNA, mỗi phân đoạn thường mã hóa cho chỉ một protein và chúng thường được tìm thấy với nhau trong một capsid. Tuy nhiên, tất cả phân đoạn không bắt buộc phải ở trong cùng một virion của virus để có thể xâm nhiễm, như được minh chứng ở virus khảm brom và vài loại virus thực vật khác.
Bộ gen virus, bất kể là loại acid nucleic nào, gần như luôn luôn có dạng sợi đơn hay sợi đôi. Bộ gen sợi đơn chứa một acid nucleic không bắt cặp, giống như một nửa của một cái thang bị cắt làm đôi ở giữa. Bộ gen sợi đôi có chứa hai acid nucleic bắt cặp bổ sung cho nhau, giống như toàn bộ cái thang. Những phần tử của một số họ virus, chẳng hạn những loại thuộc họ Hepadnaviridae, có một bộ gen mà một phần sợi đơn và một phần sợi đôi.
Với hầu hết virus có bộ gen RNA và một số với bộ gen DNA sợi đơn, những sợi đơn lẻ được cho là có tính dương (+) hoặc tính âm (-), tùy thuộc vào liệu chúng có bổ sung với RNA thông tin (mRNA) của virus hay không. RNA dương tính tức là có cùng tính với mRNA và do vậy có ít nhất một phần của nó có thể dịch mã trực tiếp bởi tế bào vật chủ. RNA âm tính thì bổ sung với mRNA và do vậy phải được chuyển thành RNA dương tính bởi một enzym RNA polymerase phụ thuộc RNA trước khi dịch mã. Danh pháp DNA với virus bộ gen ssDNA đơn tính thì tương tự như danh pháp RNA, trong đó sợi mã hóa cho mRNA của virus thì bổ sung với nó (-) và sợi không mã hóa là một bản sao của nó (+). Tuy nhiên, một vài loại virus ssDNA và ssRNA có bộ gen lưỡng tính trong đó sự phiên mã và dịch mã có thể xảy ở cả hai loại sợi trong một trung gian sao chép dạng sợi đôi. Ví dụ như geminivirus, những virus thực vật có bộ gen ssDNA, và arenavirus, những virus động vật có bộ gen ssRNA.
Kích thước bộ gen khác biệt rất lớn giữa các loài. Bộ gen virus nhỏ nhất – circovirus ssDNA, thuộc họ Circoviridae – chỉ mã hóa cho hai loại protein và có kích cỡ bộ gen vào khoảng 2 nghìn cặp base; bộ gen lớn nhất – của mimivirus – có kích thước bộ gen lên tới hơn 1,2 triệu cặp base và mã hóa cho hơn 1000 protein. Nói chung, virus RNA có bộ gen nhỏ hơn virus DNA do có tỷ lệ lỗi cao hơn khi sao chép, và có giới hạn trên về kích thước tối đa. Khi vượt qua giới hạn này, những lỗi trong bộ gen khi sao chép sẽ làm cho virus trở nên vô dụng hoặc không có tính cạnh tranh. Để khắc phục điều này, virus RNA thường có bộ gen phân đoạn – chia bộ gen thành những phân tử nhỏ hơn – để làm giảm xác suất mà một lỗi trong một bộ gen đơn phần sẽ làm mất khả năng của toàn bộ bộ gen. Trái ngược lại, virus DNA thường có bộ gen lớn hơn do độ chính xác cao của những enzym sao chép của chúng. Tuy nhiên những virus DNA sợi đơn là một ngoại lệ của quy luật này, bởi tỷ lệ đột biến ở bộ gen những loại này có thể đạt đến mức cực đại như ở trường hợp virus RNA sợi đơn.
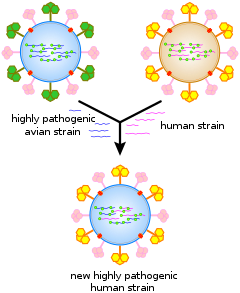
Virus cũng trải qua sự biến đổi di truyền theo một số cơ chế. Chúng bao gồm một quá trình gọi là biến động di truyền (hay trôi dạt di truyền, gentic drift) khi những base đơn lẻ trong RNA hoặc DNA đột biến thành những base khác. Hầu hết những đột biến điểm này đều "im lặng" – không thay đổi protein mà gen đó mã hóa – nhưng số khác lại có thể đem tới những lợi thế về tiến hóa như sự đề kháng với các thuốc kháng virus. Sự trôi dạt kháng nguyên diễn ra khi có một thay đổi lớn trong bộ gen của virus, có thể là hệ quả của tái tổ hợp hoặc tái sắp xếp vật liệu di truyền. Khi điều này xảy ra với virus cúm, những đại dịch có thể sẽ xảy ra. Virus RNA thường tồn tại ở dạng những loài giả (hay "biến chủng", quasispecies) hoặc những đám virus của cùng một loài nhưng có trình tự nucleoside của bộ gen hơi khác nhau. Những biến chủng này là mục tiêu chính của chọn lọc tự nhiên.
Những bộ gen phân đoạn cũng đem tới những lợi thế tiến hóa; các chủng khác nhau của cùng một virus với bộ gen phân đoạn có thể xáo trộn ngẫu nhiên và tổ hợp các gen để sản xuất ra những virus thế hệ sau (con cái) có những đặc tính độc nhất vô nhị. Điều này gọi là sự tái sắp xếp hay "sự giao phối của virus" (viral sex).
Tái tổ hợp di truyền là quá trình mà một sợi DNA bị phá vỡ và sau đó nhập vào đoạn cuối của một phân tử DNA khác. Điều này có thể xảy ra đồng thời khi virus xâm nhiễm vào tế bào và những nghiên cứu về sự tiến hóa của virus đã cho thấy rằng tái tổ hợp rất tràn lan ở các loài được nghiên cứu. Nó phổ biến ở cả virus RNA lẫn virus DNA.
Chu trình nhân lên
Những quần thể virus - do là những thực thể vô bào - nên không thể tăng trưởng thông qua sự phân chia tế bào. Thay vào đó, chúng sử dụng bộ máy và hệ trao đổi chất của tế bào vật chủ để tạo ra rất nhiều bản sao của chính chúng, và tự lắp ráp ở bên trong đó.
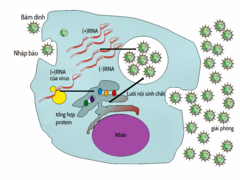
Chu trình sống của virus có sự khác nhau rất lớn giữa các loài, nhưng nhìn chung có 6 giai đoạn cơ bản trong chu trình sống (vòng đời) của virus:
- Hấp phụ là sự liên kết đặc hiệu giữa protein của vỏ capsid ở virus với những thụ thể đặc hiệu trên bề mặt tế bào vật chủ. Tính đặc hiệu này giúp xác định biên độ vật chủ của virus. Lấy ví dụ HIV, loại virus này xâm nhiễm vào một phạm vi giới hạn những tế bào bạch cầu của con người. Điều này là vì protein trên bề mặt của nó, gp120, tương tác đặc hiệu với phân tử CD4 – một thụ thể chemokine thường được tìm thấy nhiều nhất trên bề mặt của tế bào T CD4+. Cơ chế này đã tiến hóa để giúp những virus này có thể lây nhiễm chỉ trên những tế bào mà chúng có khả năng nhân lên trong đó. Sự hấp phụ (bám lấy) thụ thể có thể tạo ra những thay đổi đối với những protein của vỏ bọc virus, dẫn đến sự hợp nhất lớp lipid kép của màng tế bào và màng virus, hoặc tạo ra những biến đổi của protein bề mặt của virus (khi không có vỏ bọc) để cho phép virus đi được vào bên trong.
- Xâm nhập sau khi đã hấp phụ: Virion đi vào trong tế bào vật chủ nhờ sự nhập bào qua trung gian thụ thể hoặc sự hợp nhất màng. Sự xâm nhiễm của virus lên tế bào thực vật và nấm khác biệt với sự xâm nhiễm lên tế bào động vật. Thực vật có một thành tế bào rắn chắc được làm từ cellulose, và nấm cũng có thành tế bào từ chitin, cho nên hầu hết virus đều chỉ có thể đi vào trong những tế bào này sau một tổn thương nào đấy lên thành tế bào. Tuy nhiên, gần như tất cả virus (như virus khám thuốc lá) lại có thể di chuyển trực tiếp từ tế bào này qua tế bào khác, dưới dạng những phức hợp nucleoprotein sợi đơn thông qua các lỗ gọi là kênh liên bào (plasmodesma). Vi khuẩn, giống với thực vật, cũng có một thành tế bào rất mạnh mẽ mà virus buộc phải chọc thủng để xâm nhiễm vào tế bào. Dù vậy, bởi thành tế bào vi khuẩn kém dày hơn nhiều so với thành tế bào thực vật do kích thước nhỏ hơn rất nhiều của chúng, nên một số virus đã phát triển những cơ chế giúp bơm bộ gen của chúng vào tế bào vi khuẩn xuyên qua thành tế bào, trong khi vỏ capsid vẫn còn ở bên ngoài.
- Lột vỏ là quá trình mà vỏ capsid của virus bị loại bỏ: nó có thể bị tan rã do enzym của virus hoặc tế bào chủ hay bởi sự phân ly đơn giản; và kết quả cuối cùng là sự giải phóng ra acid nucleic của bộ gen virus.
- Sự nhân lên của virus chủ yếu liên quan đến sự nhân lên của bộ gen virus. Quá trình nhân lên bao gồm sự tổng hợp những RNA thông tin (mRNA) của virus từ những gen "sớm" (có ngoại lệ là virus RNA dương tính), sự tổng hợp protein, việc lắp ráp của những protein có thể có, và sau cùng là sự sao chép bộ gen virus được quy hoạch bởi biểu hiện của protein "sớm" hoặc protein điều hòa. Việc này có thể kéo theo một hoặc nhiều chu kỳ tổng hợp mRNA - đối với những virus phức tạp có bộ gen lớn - tạo ra sự biểu hiện gen "muộn" của protein virion hoặc protein cấu trúc.
- Sau sự tự lắp ráp do cấu trúc quy định để tạo nên các phần tử virus, thì thường xảy ra một số thay đổi trong các protein. Ở những virus như HIV, sự thay đổi này (đôi khi được gọi là 'sự thành thục') diễn ra sau khi virus đã được giải phóng ra khỏi tế bào chủ.
- Virus có thể được giải phóng ra khỏi tế bào vật chủ nhờ tiêu bào (hay làm tan tế bào), một quá trình tiêu diệt tế bào bằng cách phá tan màng tế bào (và thành tế bào nếu có) của vật chủ. Đây là một đặc tính của nhiều virus động vật và vi khuẩn. Một số virus trải qua chu trình tiềm tan, khi mà bộ gen của virus được kết hợp do tái tổ hơp di truyền vào một vị trí cụ thể trong chromosome của vật chủ. Bộ gen của virus lúc này được gọi là một "provirus", hoặc trong trường hợp của bacteriophages là một "prophage". Bất cứ khi nào vật chủ phân chia, bộ gen của virus cũng được nhân lên cùng. Bộ gen này phần lớn thời gian sẽ không hoạt động ở bên trong vật chủ, tuy nhiên, vào một thời điểm nào đó, provirus hay prophage có thể làm hoạt hóa virus, điều này có thể sẽ làm tan tế bào vật chủ. Những virus có vỏ bọc (ví dụ HIV) có đặc trưng là giải phóng khỏi tế bào chủ nhờ "nảy chồi". Trong quá trình này, virus sẽ nhận được vỏ bọc của chúng, là một phần nhỏ đã thay đổi do màng sinh chất hoặc các lưới nội chất khác của tế bào vật chủ.
Vật liệu di truyền bên trong, và phương pháp nhân lên của những vật liệu đó, khác nhau đáng kể giữa những loại virus khác nhau.
- Virus DNA
- Quá trình sao chép bộ gen ở hầu hết virus DNA diễn ra trong nhân tế bào. Nếu tế bào có thụ thể thích hợp trên bề mặt của nó, những virus này đôi khi sẽ đi vào trong tế bào bằng cách hợp nhất ngay với màng tế bào (ví dụ virus herpes) hoặc – hoặc thông thường hơn là bằng nhập vào qua trung gian thụ thể. Hầu hết virus DNA đều hoàn toàn phụ thuộc vào bộ máy sinh tổng hợp DNA và RNA, và bộ máy xử lý RNA của tế bào chủ; tuy nhiên, virus với bộ gen lớn hơn có thể tự mã hóa phần lớn bộ máy này. Ở sinh vật nhân chuẩn, bộ gen của virus phải xuyên qua màng nhân để tiếp cận với bộ máy trên, trong khi ở vi khuẩn, chúng chỉ cần đi vào bên trong tế bào là có thể tiếp cận.
- Virus RNA
- Sự sao chép thường xuyên diễn ra ở tế bào chất. Virus RNA có thể được chia làm một trong bốn nhóm khác nhau phụ thuộc vào phương thức sao chép của chúng. Sự phân cực (tức khả năng nó có thể được ribosome sử dụng trực tiếp bởi ribosome để tạo ra protein hay không) của viru RNA sợi đơn là một tiêu chí quan trọng guyết định cơ chế sao chép; trong khi tiêu chí khác đó là vật liệu di truyền là sợi đơn hay sợi đôi. Tất cả virus RNA đều sử dụng enzym sao chép RNA của chính chúng để tạo ra những bản sao của bộ gen.
- Virus phiên mã ngược
- Những loại virus này có thể là virus RNA sợi đơn, ssRNA, (Retroviridae, Metaviridae, Pseudoviridae) hoặc DNA sợi đôi, dsDNA, (Caulimoviridae và Hepadnaviridae). Những virus phiên mã ngược với bộ gen RNA (retrovirus), sử dụng một DNA trung gian cho quá trình sao chép, trong khi những loại có bộ gen DNA (pararetrovirus) sử dụng trung gian là RNA. Cả hai loại đều có enzym phiên mã ngược (reverse transcriptase), hoặc enzym DNA polymerase phụ thuộc vào RNA, để thực hiện việc chuyển đổi acid nucleic. Retrovirus sẽ tích hợp DNA tạo ra từ quá trình phiên mã ngược vào bộ gen vật chủ như một provirus (virus tiềm tan) như một bước trong chu trình nhân lên; còn pararetrovirus thì lại không làm vậy, dù bản sao bộ gen được tích hợp của một số loại pararetrovirus đặc biệt cũng đã được tìm thấy. Nhìn chung những retrovirus và pararetrovirus nhạy cảm với các loại thuốc kháng virus có khả năng ức chế enzym phiên mã ngược, ví dụ như zidovudinehay lamivudine. Một ví dụ của retrovirus là HIV, và ví dụ cho pararetrovirus là họ Hepadnaviridae, trong đó có virus viêm gan B.
Ảnh hưởng đến tế bào vật chủ
Phạm vi của những tác động về cấu trúc và sinh hóa của virus lên tế bào vật chủ là rất rộng lớn. Chúng được gọi là những tác động bệnh tích tế bào. Hầu hết những sự xâm nhiễm virus cuối cùng đều dẫn đến cái chết của tế bào vật chủ. Nguyên nhân gây ra chết bao gồm làm tan tế bào (tiêu bào), thay đổi màng ngoài tế bào và chết rụng tế bào. Thường tế bào bị chết là do việc ngừng lại các hoạt động thông thường của nó gây ra do sự ức chế của những protein đặc trưng của virus, dù không phải tất cả số đó đều là thành phần của các hạt virus.
Một số virus không gây ra sự thay đổi rõ ràng đối với tế bào bị nhiễm. Tế bào mà virus tiềm tan và bất hoạt thì cho thấy rất ít dấu hiệu bị nhiễm và thường hoạt động bình thường. Điều này gây nên sự nhiễm bệnh dai dẳng và virus thường 'ngủ đông' trong nhiều tháng hoặc nhiều năm. Điển hình cho việc này là những virus herpes. Một số loại, như virus Epstein-Barr, có thể khiến tế bào tăng sinh mà không gây ra khối u ác tính, trong khi một số loại khác như virus papilloma lại được xem là một nguyên nhân được xác nhận của bệnh ung thư.
Biên độ vật chủ
Virus chính là những thực thể sinh học dồi dào nhất từ trước tới nay trên Trái Đất, và chúng có số lượng đông hơn tất cả những loài khác cộng lại với nhau. Chúng có thể xâm nhiễm lên tất cả các dạng tế bào bao gồm của động vật, thực vật, vi khuẩn, vi khuẩn cổ và nấm. Tuy nhiên, những loại khác nhau của virus thì chỉ có thể lây nhiễm trên một phạm vi giới hạn những vật chủ khác nhau, và nhiều loại có tính đặc hiệu loài. Một số, ví dụ như virus đậu mùa, chỉ có thể nhiễm vào một loài duy nhất – trong trường hợp này là con người, và vì thế chúng được nói rằng có "biên độ vật chủ" (host range) hẹp. Những virus khác, ví dụ như virus dại, có thể lây lan trên nhiều loài động vật có vú, và do vậy có biên độ rộng. Virus mà chỉ lây nhiễm vào thực vật thì vô hại với động vật, và hầu hết virus mà xâm nhiễm lên các động vật khác thì vô hại với con người. Biên độ vật chủ của một số virus vi khuẩn - bacteriophage - thì bị giới hạn vào chỉ duy nhất một chủng vi khuẩn và chúng có thể được dùng để truy nguyên nguồn gốc bùng phát bệnh truyền nhiễm bằng một phương pháp gọi là phân loại bằng phage (phage typing).
Phân loại
Phân loại virus tìm cách mô tả tính đa dạng của virus bằng cách đặt tên và nhóm chúng dựa trên cơ sở là những điểm tương đồng. Năm 1962, André Lwoff, Robert Horne, và Paul Tournier là những người đầu tiên phát triển một phương pháp phân loại virus, dựa trên hệ thống thứ bậc của Linnaeus. Hệ thống này tạo cơ sở cho sự phân loại theo ngành, lớp, bộ, họ, chi và loài. Virus được nhóm lại dựa theo những thuộc tính chung của chúng (mà không phải của vật chủ) và loại acid nucleic tạo nên bộ gen. Về sau, Ủy ban Quốc tế về Phân loại Virus được thành lập. Tuy nhiên, virus không được phân loại theo ngành hay lớp, do bộ gen có kích thước nhỏ và tỷ lệ đột biến cao khiến chúng khó xác định tổ tiên ở trên cấp Bộ. Vì vậy, hệ thống phân loại Baltimore đã được sử dụng để bổ sung thay thế cho các cấp bậc truyền thống này.
Hệ thống phân loại ICTV
Ủy ban Quốc tế về Phân loại Virus (ICTV) đã phát triển hệ thống phân loại hiện hành và viết ra các hướng dẫn mà đề cao hơn đến các thuộc tính nhất định của virus để duy trì tính thống nhất của chúng trong một họ. Một hệ thống phân loại thống nhất đã được thành lập. Báo cáo lần thứ 7 của ICTV đã chính thức hóa lần đầu tiên khái niệm loài virus như là taxon (nhóm) thấp nhất trong hệ thống thứ bậc phân nhánh của các nhóm virus. Tuy nhiên, hiện tại chỉ có một phần nhỏ trong sự đa dạng của virus được nghiên cứu: phân tích các mẫu lấy từ con người cho thấy rằng có khoảng 20% trình tự virus được phục hồi là chưa từng được nhìn thấy trước đó, phân tích các mẫu lấy từ môi trường, như nước biển và trầm tích đại dương, giúp phát hiện ra một lượng lớn những trình tự mà hoàn toàn mới lạ với con người.
Cấu trúc hệ thống phân loại chung như sau:
Theo bảng phân loại năm 2013 của ICTV, có sáu bộ được xác lập, bao gồm Caudovirales, Herpesvirales, Mononegavirales, Nidovirales, Picornavirales và Tymovirales. Bộ thứ bảy Ligamenvirales cũng đã được đề xuất. Ủy ban không chính thức phân biệt các phân loài, chủng và dòng phân lập. Tổng cộng hiện nay có 7 bộ, 87 họ, 19 phân họ, 349 chi và khoảng 2.827 loài
Hệ thống phân loại Baltimore

Nhà sinh vật học từng đoạt giải Nobel - David Baltimore đã phát minh ra hệ thống phân loại Baltimore. Nó được sử dụng kết hợp với hệ thống phân loại ICTV trong phân loại virus hiện đại.
Baltimore phân loại virus dựa trên cơ chế sản xuất RNA thông tin (mRNA). Virus phải tạo ra mRNA từ bộ gen của chúng để sản xuất protein và sao chép chính chúng, nhưng có những cơ chế khác nhau đã được sử dụng để đạt được điều này trong mỗi họ virus. Bộ gen virus có thể là sợi đơn (ss) hoặc sợi đôi (ds), RNA hoặc DNA, và có thể sử dụng enzym phiên mã ngược (RT) hoặc không. Thêm vào đó, những virus ssRNA có thể là dương bản (+) hoặc âm bản (−). Hệ thống phân loại này sắp xếp virus vào 7 nhóm:
- I: virus dsDNA (ví dụ Adenovirus, Herpesvirus, Poxvirus)
- II: virus ssDNA DNA (+) (ví dụ Parvovirus)
- III: virus dsRNA (ví dụ Reovirus)
- IV: virus (+)ssRNA RNA (+) (ví dụ Picornavirus, Togavirus)
- V: virus (−)ssRNA RNA (-) (ví dụ Orthomyxovirus, Rhabdovirus)
- VI: virus ssRNA-RT RNA (+) với trung gian DNA (ví dụ Retrovirus)
- VII: virus dsDNA-RT (ví dụ Hepadnavirus)
Lấy một ví dụ, virus thủy đậu, Varicella zoster virus (VZV), thuộc về bộ Herpesvirales, họ Herpesviridae, phân họ Alphaherpesvirinae, và chi Varicellovirus. VZV thuộc nhóm I trong Phân loại Baltimore do nó là một virus DNA sợi đôi và không sử dụng phiên mã ngược.
Bệnh virus ở con người

Những ví dụ về các loại bệnh thông thường ở người gây ra do virus bao gồm cảm lạnh, cúm, thủy đậu và mụn rộp ở môi. Nhiều bệnh nghiêm trọng như ebola, AIDS, cúm gia cầm, và SARS cũng gây ra bởi virus. Một số bệnh đang được điều tra xem liệu chúng cũng có một virus là tác nhân gây bệnh không, chẳng hạn mối liên hệ nếu có giữa virus herpes 6 ở người (HHV6) và các bệnh thần kinh như bệnh đa xơ cứng và hội chứng mệt mỏi mãn tính. Có những tranh cãi xung quanh bornavirus, trước thường được coi là gây nên các bệnh thần kinh ở ngựa, có thể nào là thủ phạm của những bệnh tâm thần ở người hay không.
Các virus có những cơ chế gây bệnh khác nhau ở một sinh vật, phụ thuộc phần lớn vào loài virus. Cơ chế ở cấp độ tế bào chủ yếu là tiêu bào - sự mở tan và cái chết của tế bào sau đó. Ở các sinh vật đa bào, nếu có đủ số lượng tế bào chết, toàn bộ cơ thể sẽ bắt đầu chịu tác động. Mặc dù virus cũng gây nên sự gián đoạn của cân bằng nội môi khỏe mạnh, gây nên bệnh tật, chúng có thể cùng tồn tại một cách vô hại bên trong một sinh vật. Một ví dụ là khả năng của virus herpes đơn dạng, nguyên nhân gây ra bệnh rộp môi, có thể duy trì trạng thái bất hoạt bên trong cơ thể người. Đây gọi là trạng thái tiềm ẩn (latency) và là một đặc tính của các virus herpes, bao gồm virus Epstein-Barr, gây ra bệnh sốt tuyến, và virus varicella zoster, loại gây ra thủy đậu và zona. Hầu hết loài người đều đã từng nhiễm ít nhất một trong số các loại virus herpes. Tuy nhiên, những virus tiềm ẩn này đôi khi có thể có lợi, vì sự có mặt của virus làm gia tăng sự miễn dịch chống lại các mầm bệnh vi khuẩn, ví dụ như Yersinia pestis.
Một số loại virus có thể gây ra sự nhiễm bệnh mãn tính hoặc suốt đời, khi mà virus tiếp tục nhân lên trong cơ thể bất kể có những cơ chế phòng ngự của vật chủ. Điều này rất phổ biến với bệnh do virus viêm gan B và viêm gan C. Những người bị nhiễm bệnh mãn tính gọi là những người mang (carrier), do họ đóng vai trò một nguồn dự trữ loại virus xâm nhiễm. Ở những quần thể có tỷ lệ người mang cao, căn bệnh được coi là có tính địa phương.
Dịch tễ học
Dịch tễ học virus là một nhánh của y học, nghiên cứu sự lây truyền và kiểm soát bệnh truyền nhiễm do virus gây ra ở con người. Sự lây truyền của virus có thể theo chiều dọc, tức từ mẹ sang con, hoặc chiều ngang, tức từ người này sang người khác. Những ví dụ về truyền bệnh theo chiều dọc là virus viêm gan B và HIV, ở đây trẻ con sinh ra đã bị nhiễm virus lây từ mẹ. Một ví dụ khác, hiếm hơn, là virus varicella zoster, loại mặc dù chỉ gây ra sự nhiễm bệnh tương đối nhẹ ở người, lại có thể gây tử vong với thai nhi và trẻ sơ sinh.
Truyền bệnh theo chiều ngang là cơ chế lây lan virus phổ biến nhất trong quần thể. Sự lây bệnh có thể xảy ra khi: chất dịch cơ thể được trao đổi trong hoạt động tình dục (ví dụ HIV); máu được trao đổi do truyền máu bị nhiễm bệnh hoặc dùng chung kim tiêm (ví dụ virus viêm gan C); trao đổi nước bọt qua đường miệng (ví dụ virus Epstein-Barr); thực phẩm và nước bị nhiễm bẩn đi qua đường tiêu hóa (ví dụ norovirus); hít phải các sol khí chứa virion (ví dụ virus cúm); và các vector côn trùng như muỗi đưa mầm bệnh xâm nhập qua da của người bệnh (ví dụ sốt xuất huyết). Tỷ lệ hoặc tốc độ lây truyền bệnh virus phụ thuộc vào các nhân tố như mật độ dân số, số cá thể nhạy cảm (như những người không miễn dịch), chất lượng y tế và thời tiết.
Dịch tễ học được ứng dụng để phá vỡ chuỗi lây nhiễm trong các quần thể đang phải chịu sự bùng phát dịch bệnh do virus. Các phương pháp kiểm soát được dựa trên những kiến thức về cách mà virus lây lan. Việc tìm ra nguồn bệnh của sự bùng phát dịch và việc xác định virus rất quan trọng. Một qua virus đã được định danh, chuỗi lan truyền có thể được phá vỡ nhờ các vắc-xin; hoặc khi vắc-xin không có sẵn thì vệ sinh và khử trùng cũng có thể có hiệu quả. Thông thường, những người nhiễm bệnh bị cô lập khỏi phần còn lại của cộng đồng, và những người đã tiếp xúc với virus sẽ bị đặt trong tình trạng cách ly. Để khống chế sự bùng phát bệnh lở mồm long móng ở bò tại Anh năm 2001, hàng ngàn con bò đã bị giết. Hầu hết bệnh truyền nhiễm do virus ở người và các động vật khác có thời kỳ ủ bệnh khi sự nhiễm bệnh không gây ra dấu hiệu hay triệu chứng nào. Thời gian ủ bệnh do virus dao động từ một vài ngày đến vài tuần, được biết với hầu hết bệnh do virus. Mặc dù có hơi chồng lấn lên nhau, nhưng chủ yếu diễn ra sau thời kỳ ủ bệnh là một thời kỳ lây lan— giai đoạn mà một cá thể nhiễm bệnh dễ lây nhiễm và có thể lây sang những người hay động vật khác. Điều này cũng xảy ra với nhiều bệnh virus, và hiểu biết về độ dài của hai thời kỳ rất quan trọng đối với sự kiểm soát các trận dịch. Khi sự bùng phát gây nên một tỷ lệ cao bất thường các ca bệnh trong một cộng đồng hay một vùng, chúng được gọi là những "dịch bệnh" hay "dịch" (epidemic). Nếu sự bùng phát lan ra toàn thế giới, chúng được xem như những "đại dịch" (pandemic).
Dịch và đại dịch
Những quần thể người Mỹ bản địa đã bị tàn phá bởi những căn bệnh truyền nhiễm, đặc biệt bệnh đậu mùa, do những thực dân châu Âu mang tới châu Mỹ. Người ta không rõ là đã có bao nhiêu người Mỹ bản địa bị chết do những căn bệnh ngoại lai xuất hiện sau sự có mặt của Columbus ở châu Mỹ, nhưng đã ước tính là lên tới 70% dân số bản địa. Sự hủy diệt do những căn bệnh này gây ra đã hỗ trợ đáng kể cho nỗ lực của người châu Âu để thay thế và chinh phục những cộng đồng bản địa nơi đây.
Nhìn chung, một đại dịch là một dịch bệnh mang tính toàn cầu. Đại dịch cúm 1918, diễn ra cho đến năm 1919, là một đại dịch cúm nằm trong nhóm 5 (nhóm nghiêm trọng nhất) trong Chỉ số Nghiêm trọng của Đại dịch (PSI), gây ra bởi một dạng virus cúm A nguy hiểm chết người và nghiêm trọng một cách bất thường. Các nạn nhân thường là những thanh niên khỏe mạnh, trái ngược với hầu hết các dịch cúm khác vốn chủ yếu ảnh hưởng đến trẻ em, người già, hoặc nếu không là những bệnh nhân ốm yếu. Những ước tính ban đầu nói rằng dịch cúm này đã giết chết khoảng 40–50 triệu người, trong khi những nghiên cứu gần đây hơn đề xuất rằng nó có thể đã giết hại lên tới 100 triệu người, tức khoảng 5% dân số thế giới chỉ trong năm 1918.


Nhiều nhà nghiên cứu tin rằng virus HIV có xuất xứ từ châu Phi cận Sahara trong thế kỷ XX; nó hiện nay được công nhận là một đại dịch, với ước tính khoảng 38,6 triệu người hiện sống với căn bệnh này trên toàn thế giới.Chương trình Phối hợp của Liên Hợp Quốc về HIV/AIDS (UNAIDS) và Tổ chức Y tế Thế giới (WHO) ước tính rằng AIDS đã giết hại hơn 25 triệu người tính từ khi được ghi nhận lần đầu vào 5 tháng 6 năm 1981, khiến nó là một trong những dịch bệnh hủy hoại nhất được ghi lại trong lịch sử. Chỉ trong năm 2007, có thêm khoảng 2,7 triệu ca nhiễm HIV mới và thêm 2 triệu người chết có liên quan tới HIV.
Một số mầm bệnh virus cực kỳ nguy hiểm khác là những thành viên của họ Filoviridae. Filovirus là những virus gây ra bệnh sốt xuất huyết siêu vi, bao gồm virus Ebola và virus Marburg. Virus Marburg đã thu hút sự quan tâm rộng rãi của báo chí vào tháng 4 năm 2005 với một trận bùng phát đầu tiên ở Angola. Bệnh do vi rút Ebola cũng gây ra các đợt bùng phát không liên tục với tỷ lệ tử vong cao, kể từ năm 1976 khi lần đầu tiên được xác định. Tồi tệ và gần đây nhất là dịch bệnh tại Tây Phi năm 2014.
Ngoại trừ bệnh đậu mùa, hầu hết các đại dịch đều do virus mới tiến hóa gây ra. Các virus "mới nổi" này thường là đột biến của những loại virus ít gây hại hơn đã từng lưu hành trước đây ở người hoặc động vật khác.
Hội chứng hô hấp cấp tính nặng (SARS) và hội chứng hô hấp Trung Đông (MERS) đều do các chủng coronavirus mới gây ra. Các chủng coronavirus khác được biết là gây ra các bệnh nhiễm trùng nhẹ ở người, do vậy độc lực và sự lây lan nhanh chóng của SARS—vào tháng 7 năm 2003 đã gây ra khoảng 8.000 ca mắc và 800 ca tử vong—là điều không ngờ và hầu hết các quốc gia đều không chuẩn bị kỹ càng.
Một chủng coronavirus liên quan, virus corona gây hội chứng hô hấp cấp tính nặng 2 (SARS-CoV-2), được cho là có nguồn gốc từ loài dơi, xuất hiện tại Vũ Hán, Trung Quốc vào tháng 11 năm 2019 và nhanh chóng lây lan trên toàn thế giới. Nó đã gây ra đại dịch COVID-19 bắt đầu vào năm 2020. Những hạn chế chưa từng có trong thời bình đã được đặt ra đối với việc đi lại quốc tế, và lệnh giới nghiêm đã được áp dụng ở một số thành phố lớn trên thế giới nhằm ứng phó với đại dịch.
Ung thư

Virus là một nguyên nhân được công nhận của bệnh ung thư ở con người và các loài khác. Ung thư do virus chỉ xảy ra ở một số ít người (hay động vật) bị nhiễm. Virus ung thư đến từ một loạt các họ virus, ở cả hai nhóm virus RNA và DNA, và không có bất kỳ loại đơn lẻ nào là "oncovirus" (một thuật ngữ đã lỗi thời ban đầu dùng để chỉ retrovirus chuyển đổi cấp). Sự phát triển của ung thư do virus được xác định bởi nhiều nhân tố, như sự miễn dịch của vật chủ và những đột biến bên trong vật chủ. Virus được đồng thuận là gây ra bệnh ung thư ở người bao gồm một số kiểu gen của các loại virus papilloma ở người, virus viêm gan B, virus viêm gan C, virus Epstein-Barr, virus herpes ung thư mô liên kết Kaposi và virus ưa lympho T ở người. Một loại virus ung thư ở người mới khám phá gần đây là polyomavirus (virus polyoma tế bào Merkel) mà gây ra phần lớn các ca của một dạng ung thư da hiếm gặp gọi là ung thư biểu mô tế bào Merkel. Những virus viêm gan cũng có thể phát triển thành bệnh virus mãn tính mà dẫn tới ung thư gan. Sự xâm nhiễm của virus ưa bạch cầu T ở người cũng có thể dẫn tới chứng liệt cứng chi dưới nhiệt đới và ung thư bạch cầu tế bào T trưởng thành. Virus papilloma cũng được xác định là một trong những nguyên nhân của các dạng ung thư cổ tử cung, da, hậu môn, và dương vật. Trong họ Herpesviridae, virus herpes ung thư mô liên kết Kaposi gây ra bệnh ung thư mô liên kết Kaposi (Kaposi's sarcoma) và ung thư hạch khoang cơ thể, và virus Epstein–Barr gây ra ung thư hạch Burkitt, ung thư hạch Hodgkin, rối loạn tăng sinh lympho B và ung thư vòm họng. Virus polyoma tế bào Merkel cũng có sự liên hệ mật thiết với SV40 và virus polyoma ở chuột mà đã được sử dụng như những mô hình tiêu biểu cho virus gây ung thư ở động vật trong hơn 50 năm qua.
Cơ chế phòng ngự của vật chủ
Rào chắn bảo vệ đầu tiên của cơ thể chống lại virus là hệ thống miễn dịch tự nhiên. Nó bao gồm những tế bào và các cơ chế khác giúp bảo vệ vật chủ chống lại sự xâm nhiễm một cách không đặc hiệu. Điều này có nghĩa là những tế bào của hệ thống tự nhiên sẽ ghi nhận và phản ức với mầm bệnh theo một cách chung chung, nhưng không giống với hệ thống miễn dịch thích ứng, nó không cung cấp sự miễn dịch bảo vệ hoặc lâu dài với vật chủ.
Can thiệp RNA là một cơ chế phòng ngự bẩm sinh khác chống lại virus. Nhiều loại virus có chiến lược nhân lên mà có sự tham gia của RNA sợi đôi (dsRNA). Khi một virus như vậy lây nhiễm vào tế bào, nó giải phóng phân tử RNA của nó, thứ mà ngay lập tức sẽ gắn vào một phức hợp protein gọi là dicer, một enzym cắt RNA thành những mảnh nhỏ. Một con đường sinh hóa gọi là phức hợp RISC được kích hoạt, giúp phân hủy mRNA của virus và tế bào sẽ sống sót qua khỏi sự nhiễm bệnh. Rotavirus tránh cơ chế này bằng cách không lột vỏ hoàn toàn bên trong tế bào, và giải phóng mRNA mới qua các lỗ trong vỏ capsid trong của hạt virus. Bộ gen dsRNA nhờ thế vẫn được bảo vệ bên trong hạch tâm của virion.
Khi một hệ thống miễn dịch thích ứng của một động vật có xương sống gặp phải một virus, nó sẽ sản sinh ra các kháng thể đặc hiệu, gắn vào virus và thường làm nó không nhiễm bệnh. Đây gọi là miễn dịch dịch thể. Có hai loại kháng thể rất quan trọng, Loại đầu tiên gọi là IgM, rất hiệu quả trong việc trung hòa virus, được sản xuất bởi tế bào của hệ miền dịch chỉ trong vài tuần. Loại thứ hai, gọi là IgG, được sản xuất không giới hạn. Sự hiện diện của IgM trong máu vật chủ được sử dụng để kiểm tra sự nhiễm bệnh cấp tính, trong khi IgG biểu thị về sự nhiễm bệnh vào một lúc nào đó trong quá khứ. Kháng thể IgG cũng được đo trong các xét nghiệm miễn dịch.
Kháng thể cũng tiếp tục trở thành một cơ chế phòng ngự hữu hiệu ngay cả sau khi virus đã khống chế và xâm nhập vào tế bào chủ. Một protein trong tế bào này, gọi là TRIM21, có thể gắn vào kháng thể trên bề mặt của phần tử virus. Điều này chuẩn bị cho sự phá hủy virus kế tiếp thực hiện bởi các enzym của hệ thống proteosome trong tế bào.
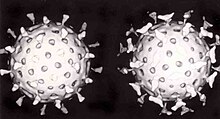
Sự bảo vệ thứ hai của động vật có xương sống chống lại virus là miễn dịch qua trung gian tế bào, liên quan đến các tế bào miễn dịch được biết đến với tên tế bào T. Tế bào của cơ thể liên tục hiển thị các đoạn protein ngắn trên bề mặt tế bào, và nếu tế bào T nhận ra được một phân đoạn được nghi ngờ là của virus ở đây, tế bào chủ sẽ bị các tế bào giết tự nhiên T tiêu diệt, và những tế bào T đặc hiệu với virus sẽ sinh sôi nhanh chóng. Những tế bào như đại thực bào là những chuyên gia trong quá trình trình diện kháng nguyên. Sự sản xuất ra các interferon cũng là một cơ chế bảo vệ quan trọng. Đây là một hormone do cơ thể sinh ra khi virus xuất hiện. Vai trò của nó trong sự miễn dịch rất phức tạp; nó cuối cùng sẽ làm dừng lại sự sinh sản của virus bằng cách giết chết tế bào bị nhiễm và các tế bào xung quanh nó.
Không phải tất cả sự xâm nhiễm virus đều tạo ra một phản ứng miễn dịch bảo vệ theo cách trên. HIV trốn tránh khỏi hệ thống miễn dịch bằng cách liên tục thay đổi trình tự amino acid của protein trên bề mặt virion. Loại virus dai dẳng này trốn tránh sự kiểm soát miễn dịch bằng cách cô lập, phong tỏa trình diện kháng nguyên, đề kháng lại cytokine, tránh khỏi hoạt động của tế bào giết tự nhiên và chết rụng tế bào, cùng sự trôi dạt kháng nguyên. Một số loại virus khác, được gọi là virus hướng thần kinh, có thể lây lan đến các tế bào thần kinh nơi hệ thống miễn dịch không thể nào tiếp cận chúng.
Phòng tránh và chữa trị
Vì lý do virus sử dụng các con đường trao đổi chất quan trọng trong tế bào cho việc sinh sản, nên nói chung rất khó để loại bỏ chúng mà không sử dụng những loại thuốc gây ra những ảnh hưởng độc hại đến tế bào chủ. Những cách tiếp cận y tế hiệu quả nhất đối với bệnh do virus là sử dụng tiêm chủng để cung cấp khả năng miễn dịch đối với sự xâm nhiễm của virus, và dùng thuốc kháng virus để can thiệp có chọn lọc lên quá trình nhân lên của chúng.
Vắc-xin
Tiêm chủng là một biện pháp có phi tổn thấp và hữu hiệu để ngăn chặn sự xâm nhiễm của virus. Các vắc-xin đã được sử dụng để ngăn chặn để ngăn ngừa virus lây nhiễm trong một thời gian dài trước khi tìm thấy những loại virus thực sự. Việc sử dụng vắc-xin đã dẫn tới sự suy giảm đáng kể trong tỷ lệ mắc bệnh (bị ốm) và tỉ lệ tử vong (bị chết) bởi các bệnh liên quan tới virus như bại liệt, sởi, quai bị và rubella. Đặc biệt bệnh đậu mùa đã hoàn toàn bị loại trừ nhờ những chiến dịch tiêm chủng quy mô lớn. Vắc-xin có khả năng ngăn chặn hơn 13 bệnh lây nhiễm do virus khác ở người, và nhiều bệnh hơn ở động vật. Vắc-xin có thể bao gồm những virus còn sống đã làm giảm độc lực hoặc virus đã chết, hay những protein của virus (kháng nguyên). Vắc-xin sống có chứa các dạng đã bị làm suy yếu của virus, những dạng này không gây ra bệnh nhưng lại có thể tạo ra sự miễn dịch. Vắc-xin sống có thể gây nguy hiểm nếu được đưa vào cơ thể người mà có hệ miễn dịch yếu (người bị suy giảm miễn dịch), bởi ở những người này, virus suy yếu vẫn có thể gây ra bệnh như ban đầu. Công nghệ sinh học và các kĩ thuật di truyền đã được sử dụng để sản xuất ra vắc-xin tiểu đơn vị. Loại vắc-xin này chỉ sử dụng duy nhất protein ở vỏ capsid của virus. Vắc-xin virus viêm gan B là ví dụ cho loại vắc-xin này. Vắc-xin tiểu đơn vị thì an toàn với những bệnh nhân bị suy giảm miễn dịch do chúng không thể nào gây ra căn bệnh. Vắc-xin virus sốt vàng, một chủng vắc-xin sống đã giảm độc lực được gọi là 17D, được xem có thể là chủng vắc-xin an toàn và hiệu quả nhất mà con người từng tạo ra.
Thuốc kháng virus

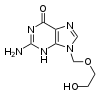
Những loại thuốc kháng virus thường là những chất tương tự nucleoside - thành phần xây dựng nên DNA - khiến virus nhầm lẫn và tích hợp chúng vào trong bộ gen của virus lúc đang ở trong quá trình sinh sản. Vòng đời của virus sau đó sẽ bị dừng lại do DNA mới tổng hợp rơi vào trạng thái bất hoạt. Điều này là bởi những chất tương tự này thiếu đi nhóm hydroxyl (-OH), nhóm mà cùng với các nguyên tử phosphor, liên kết với nhau để tạo nên một bộ "xương sống" mạnh mẽ cho phân tử DNA. Việc làm này gọi là sự ngắt mạch DNA. Những ví dụ cho chất tương tự nucleoside là aciclovir, thuốc kháng lây nhiễm virus Herpes đơn dạng, và lamivudine, thuốc kháng lây nhiễm HIV và HBV. Aciclovir chính là một trong những loại thuốc kháng virus lâu đời nhất và thường được kê đơn nhất. Những loại thuốc kháng virus khác hiện được sử dụng hướng tới những giai đoạn khác nhau trong chu kỳ sống của virus. Với HIV, loại virus phụ thuộc vào một enzym phân giải protein có tên HIV-1 protease để có thể hoàn toàn xâm nhiễm, thì có rất nhiều loại thuốc được gọi với tên chung là thuốc ức chế protease có vai trò bất hoạt loại enzym này.
Bệnh viêm gan C là một bệnh gây ra bởi một virus RNA. Ở 80% người bị nhiễm, căn bệnh này là mãn tính, và nếu không được điều trị thì người nhiễm sẽ mắc bệnh trong suốt phần đời còn lại của họ. Tuy nhiên, có một cách điều trị hiệu quả đó là sử dụng thuốc tương tự ucleoside ribavirin kết hợp với interferon. Việc điều trị những người mang virus viêm gan B mãn tính bằng cách sử dụng chiến thuật tương tự với thuốc lamivudine cũng đã được phát triển.
Lây nhiễm trên các loài khác
Virus lây nhiễm lên tất cả các dạng sống tế bào và, dù virus xuất hiện rất phổ biến, thì mỗi loài sinh vật có tế bào đều có một phạm vi cụ thể những virus mà thường chỉ xâm nhiễm duy nhất lên loài đó. Một số virus, gọi là những vệ tinh, chỉ có nhân lên ở trong những tế bào mà đã từng bị xâm nhiễm bởi một virus khác. Virus là những tác nhân gây bệnh quan trọng ở gia súc. Những bệnh như lở mồm long móng và lưỡi xanh là những ví dụ về bệnh gia súc gây ra do virus. Các loài vật cưng như chó, mèo hay ngựa, nếu không được tiêm phòng, cũng dễ nhạy cảm với những bệnh virus nghiêm trọng. Bệnh virus parvo ở chó gây ra do một loại virus DNA nhỏ và sự nhiễm bệnh thường dẫn đến tử vong ở chó con. Các loài không xương sống, ví dụ như ong mật, cũng dễ bị lây nhiễm nhiều bệnh do virus. Tuy nhiên, hầu hết virus đều chung sống một cách vô hại với vật chủ và không gây ra bất kỳ dấu hiệu hay triệu chứng bệnh nào.
Thực vật

Có rất nhiều loại virus thực vật, nhưng chúng thường chỉ gây ra tổn thất về sản lượng, và sẽ không khả thi về mặt kinh tế khi cố gắng kiểm soát chúng. Virus thực vật thường lây lan từ cây này sang cây khác qua những sinh vật trung gian, gọi là các vector. Chúng thường là côn trùng, nhưng một số loài nấm, giun tròn và sinh vật đơn bào cũng được chứng minh là những vector. Khi sự kiểm soát các bệnh virus thực vật được công nhận là kinh tế, ví dụ ở các cây ăn quả lâu năm, người ta thường tập trung các nỗ lực vào việc giết chết các vector và loại bỏ những vật chủ thay thế như cỏ dại. Virus thực vật không thể lây nhiễm lên con người và các loài động vật khác.
Thực vật có những cơ chế bảo vệ hiệu quả và phức tạp chống lại virus. Một trong những cơ chế hiệu quả nhất là sự có mặt của những gen gọi là gen đề kháng (R). Mỗi gen R giúp đề kháng một loại virus cụ thể bằng cách kích hoạt cái chết của những khu vực cục bộ các tế bào xung quanh tế bào bị nhiễm, những vùng này có thể nhìn thấy bằng mắt thường dưới dạng những đốm lớn. Điều này sẽ ngăn ngừa căn bệnh lây lan.Can thiệp RNA cũng là một cách phòng ngự hiệu quả ở thực vật. Khi chúng bị nhiễm bệnh, thực vật thường tạo ra những chất khử trùng tự nhiên để tiêu diệt virus, chẳng hạn như acid salicylic, nitơ monoxit và các phân tử oxy phản ứng.
Những phần tử virus hoặc giống virus (virus-like particle, VLP) ở thực vật có những ứng dụng trong cả công nghệ sinh học và công nghệ nano. Vỏ capsid của phần lớn virus thực vật là những cấu trúc đơn giản và mạnh mẽ, có thể được sản xuất với số lượng lớn bởi bệnh lây nhiễm ở thực vật hay bởi biểu hiện của một loạt các hệ thống dị thể. Các phần tử virus thực vật có thể được chỉnh sửa về cả di truyền và hóa học để 'đóng gói' các vật liệu ngoại lai, và có thể tích hợp vào trong các cấu trúc siêu phân tử để sử dụng trong công nghệ sinh học.
Vi khuẩn

Bacteriophage là một nhóm virus phổ biến và đa dạng, và là dạng thực thể sinh học nhiều nhất trong môi trường nước – chúng có số lượng nhiều gấp tới 10 lần số vi khuẩn ở các đại dương, đạt mật độ khoảng 250.000.000 bacteriophage mỗi mililít nước biển. Những virus này xâm nhiễm đặc hiệu vào các vi khuẩn bằng cách liên kết với những thụ thể trên bề mặt và sau đó đi vào trong tế bào. Trong một khoảng thời gian ngắn, đôi khi chỉ vài phút, polymerase của vi khuẩn sẽ bắt đầu dịch mã mRNA của virus thành protein. Những protein này sẽ tiếp tục trở thành những virion mới bên trong tế bào, hoặc các protein hỗ trợ, giúp đỡ lắp ráp virion mới, hay những protein tham gia vào tiêu bào. Những enzym virus xúc tác cho sự phá vỡ màng tế bào, và trong trường hợp của phage T4, thì chỉ hơn 20 phút sau khi bơm vào có tới hơn 300 phage có thể được giải phóng.
Cách thức chính mà vi khuẩn bảo vệ bản thân chúng khỏi bacteriophage là sản sinh ra những enzym mà phá hủy DNA ngoại lai. Những enzym này gọi là endonuclease giới hạn, cắt đi DNA mà bacteriophage tiêm vào trong tế bào vi khuẩn. Vi khuẩn cũng chứa một hệ thống có sử dụng các trình tự CRISPR để giữ lại các mảnh của bộ gen virus mà vi khuẩn đã từng tiếp xúc trong quá khứ, điều cho phép chúng ngăn chặn sự sao chép của virus thông qua một dạng can thiệp RNA. Hệ thống di truyền này sẽ cung cấp cho vi khuẩn sự miễn dịch thu được với sự xâm nhiễm của virus.
Cổ khuẩn
Một số loại virus sinh sản được bên trong cổ khuẩn; chúng là những virus DNA sợi đôi với hình dáng bất thường và đôi khi là độc nhất vô nhị. Những virus này đã được nghiên cứu chi tiết nhất ở những cổ khuẩn ưa nhiệt, đặc biệt trong bộ Sulfolobales và Thermoproteales. Sự phòng ngự chống lại những virus này có thể liên quan tới sự can thiệp RNA từ các trình tự DNA lặp lại bên trong bộ gen cổ khuẩn, thứ mà có liên hệ với những gen của virus.
Vai trò trong hệ sinh thái nước
Một muỗng cà phê nước biển có chứa khoảng một triệu hạt virus. Chúng rất cần thiết cho sự điều hòa hệ sinh thái nước mặn và nước ngọt. Hầu hết những virus này đều là bacteriophage, những loại vô hại đối với thực vật và động vật. Chúng lây nhiễm và tiêu diệt vi khuẩn trong các cộng đồng vi sinh dưới nước, làm nên cơ chế quan trọng nhất trong sự tái chế carbon ở môi trường đại dương. Những phân tử hữu cơ được giải phóng ra khỏi tế bào vi khuẩn sẽ kích thích sự phát triển mới của vi khuẩn và tảo.
Những vi sinh vật chiếm hơn 90% tổng sinh khối ở biển. Ước tính rằng virus đã giết và giải phóng xấp xỉ 20% của lượng sinh khối đấy mỗi ngày và chúng có số lượng nhiều gấp 15 lần số lượng của vi khuẩn và cổ khuẩn ở đại dương. Virus là tác nhân chính gây nên sự tàn phá nhanh chóng của các đợt tảo nở hoa, hiện tượng mà thường giết hại những sinh vật biển khác. Số lượng virus trong đại dương giảm khi đi ra ngoài khơi và sâu vào trong nước, nơi có ít vật chủ hơn.
Những ảnh hưởng của virus đại dương rất sâu rộng; bằng cách gia tăng lượng quang hợp ở biển, virus là nguyên nhân gián tiếp làm giảm lượng khí carbon dioxide trong khí quyển vào khoảng xấp xỉ 3 tỉ tấn cacbon mỗi năm.
Giống như bất cứ sinh vật nào, động vật có vú ở biển cũng nhạy cảm với các bệnh do virus. Vào năm 1988 và 2001, hàng ngàn con hải cẩu đã bị giết hại ở châu Âu do virus sài sốt hải cẩu (PDV). Nhiều loại virus khác, như calicivirus, herpesvirus, adenovirus và parvovirus, cũng lưu hành trong cách quần thể động vật có vú đại dương.
Vai trò trong tiến hóa
Virus là một phương tiện tự nhiên quan trọng để chuyển giao các gen giữa những loài khác nhau, điều góp phần gia tăng đa dạng di truyền và tạo ra sự tiến hóa. Có quan niệm rằng virus đóng một vai trò chủ chốt trong sự tiến hóa sơ khai, trước khi có sự đa dạng của vi khuẩn, vi khuẩn cổ và sinh vật nhân chuẩn và vào giai đoạn của tổ tiên chung cuối cùng của sự sống trên Trái Đất. Cho đến ngày nay, virus vẫn là một trong những nguồn dự trữ đa dạng di truyền lớn nhất mà chưa được khám phá trên Trái Đất.
Ứng dụng
Khoa học sự sống và y học

Virus là những sinh vật rất quan trọng trong nghiên cứu sinh học phân tử và sinh học tế bào do chúng cung cấp những hệ thống đơn giản mà có thể sử dụng để thao tác và nghiên cứu các chức năng của tế bào. Những nghiên cứu và ứng dụng của virus đã đem tới những thông tin giá trị về các khía cạnh của sinh học tế bào. Ví dụ, virus rất hữu dụng trong nghiên cứu di truyền học và giúp đỡ cho sự hiểu biết của chúng ta về các cơ chế cơ bản của di truyền học phân tử, như tái bản DNA, phiên mã, xử lý RNA, dịch mã, vận chuyển protein, và miễn dịch.
Di truyền học thường sử dụng virus như những vector để đưa các gen vào tế bào mà họ đang nghiên cứu. Điều này rất có ích để tạo nên tế bào mà sản xuất ra một chất ngoại lai, hoặc nghiên cứu ảnh hưởng của việc đưa gen mới vào trong bộ gen. Theo cách tương tự, liệu pháp virus (virotherapy) sử dụng những virus như những vector để điều trị các bệnh khác nhau, do chúng có thể nhắm đến các tế bào và DNA một cách đặc hiệu. Điều này cho thấy ứng dụng rất triển vọng của virus trong việc điều trị bệnh ung thư và trong liệu pháp gen. Các nhà khoa học Đông Âu cũng đã sử dụng liệu pháp phage như là một sự thay thế cho thuốc kháng sinh trong một thời gian, và mối quan tâm đến cách tiếp cận này đang gia tăng, bởi vì sự đề kháng kháng sinh ở mức độ cao hiện nay đã được tìm thấy ở một số vi khuẩn gây bệnh. Sự biểu hiện các protein dị thể bởi virus là cơ sở cho một vài quy trình sản xuất hiện đang được dùng để sản xuất nên nhiều protein như kháng nguyên của vắc-xin và kháng thể. Người ta gần đây cũng đã phát triển những quy trình công nghiệp sử dụng vector virus; và một số các protein dược phẩm hiện nay đang trong quá trình thử nghiệm tiền lâm sàng và lâm sàng.
Khoa học vật liệu và công nghệ nano
Những xu hướng hiện nay trong công nghệ nano hứa hẹn sẽ đem lại việc ứng dụng virus một cách linh hoạt hơn. Theo quan điểm của các nhà khoa học vật liệu, virus có thể được xem như những hạt nano hữu cơ. Bề mặt của chúng chứa những công cụ đặc biệt được thiết kế để vượt qua các rào chắn của tế bào vật chủ. Kích cỡ và hình dạng của virus, cũng như số lượng và bản chất của các nhóm chức năng trên bề mặt chúng, đã được xác định một cách chính xác. Như vậy, virus có thể được sử dụng phổ biến trong khoa học vật liệu như giá đỡ cho những sửa đổi bề mặt được liên kết cộng hóa trị. Một phẩm chất đặc biệt của virus là chúng có thể được điều chỉnh nhờ tiến hóa có định hướng. Các kĩ thuật mạnh mẽ được phát triển trong các ngành khoa học sự sống đã đang trở thành nền tảng cho hướng tiếp cận kĩ thuật về vật liệu nano, mở ra một loạt những ứng dụng rộng rãi vượt ra khỏi sinh học và y học.
Do kích thước, hình dạng, và cấu trúc hóa học dễ xác định, virus được sử dụng như những bản mẫu để tạo nên những vật liệu ở cấp độ nano. Những ví dụ gần đây bao gồm nghiên cứu tại Phòng thí nghiệm Nghiên cứu Hải quân Mỹ ở Washington, D.C., sử dụng các phần tử virus khảm đậu đũa (CPMV) để khuếch đại những tín hiệu trong cảm biến dựa trên DNA microarray. Trong ứng dụng này, các hạt virus tách riêng các thuốc nhuộm huỳnh quang sử dụng cho báo hiệu để ngăn ngừa sự hình thành các dimer (chất nhị trùng) mà đóng vai trò là những tác nhân dập tắt huỳnh quang. Một ví dụ khác là việc sử dụng CPMV như một bộ cắm dây có kích thước nano dùng trong điện tử học phân tử.
Virus nhân tạo
Nhiều virus có thể được tổng hợp "từ đầu" ("de novo") và virus nhân tạo (virus tổng hợp) đầu tiên đã được tạo ra năm 2002. Mặc dù phần nào bị hiểu sai, nó thực chất không phải là một virus thực sự được tổng hợp, mà thay vào đó là bộ gen DNA của nó (nếu đó là virus DNA) hoặc một bản sao DNA bổ sung (cDNA) của bộ gen của nó (nếu đó là virus RNA). Với nhiều họ virus, DNA hay RNA nhân tạo dạng trần (khi được chuyển đổi lại bằng enzym từ phân tử cDNA) có khả năng lây nhiễm khi được đưa vào bên trong tế bào. Chúng có chứa tất cả các thông tin cần thiết để sản xuất những virus mới. Công nghệ này hiện nay được sử dụng để nghiên cứu những chiến lược vắc-xin mới. Khả năng tổng hợp nên những virus đã có những hệ quả sâu rộng,bởi virus nhờ vậy không bao giờ có thể coi là tuyệt chủng nếu trình tự bộ gen của chúng còn được biết đến và các tế bào cho phép tiếp nhận vẫn có sẵn. Hiện nay, trình tự bộ gen đầy đủ của 2408 loại virus khác nhau (bao gồm cả đậu mùa) đã được công bố công khai ở một cơ sở dữ liệu trực tiếp, được duy trì bởi Viện Y tế Quốc gia Hoa Kỳ.
Vũ khí sinh học
Khả năng tạo nên những dịch bệnh trong xã hội loài người của virus đã đưa tới những lo ngại rằng virus có thể được vũ khí hóa cho chiến tranh sinh học. Mối lo lắng này được nâng lên bởi sự tái tạo thành công loại virus cúm Tây Ban Nha năm 1918 nổi tiếng ở một phòng thí nghiệm. Virus đậu mùa cũng đã tàn phá nhiều xã hội trong suốt lịch sử loài người trước khi nó bị tiêu diệt hoàn toàn. Chỉ có duy nhất hai trung tâm chính thức lưu trữ virus bệnh đậu mùa – Phòng thí nghiệm Vector của Nga và Trung tâm Kiểm soát Bệnh của Hoa Kỳ. Tuy nhiên, nỗi sợ rằng nó có thể được sử dụng làm vũ khí không hẳn là không có cơ sở; vắc-xin đậu mùa đôi lúc có những tác dụng phụ nghiêm trọng – trong những năm sau cùng trước khi bệnh đậu mùa bị tiêu diệt, nhiều người đã bị bệnh nghiêm trọng do hậu quả của tiêm chủng còn nhiều hơn số người bị nhiễm trực tiếp. Việc tiêm phòng đậu mùa cũng đã không còn được áp dụng rộng rãi, và như vậy, phần lớn dân số loài người hiện đại đều gần như không có sự đề kháng nào được thiết lập chống lại bệnh đậu mùa.
Tham khảo
Tài liệu
- Collier, Leslie; Balows, Albert; Sussman, Max (1998) Topley and Wilson's Microbiology and Microbial Infections ninth edition, Volume 1, Virology, volume editors: Mahy, Brian and Collier, Leslie. Arnold. ISBN 0-340-66316-2, doi:10.1002/9780470688618; ISBN 0-340-614706 doi:10.1056/NEJM199810293391820
- Dimmock, N.J; Easton, Andrew J; Leppard, Keith (2007) Introduction to Modern Virology sixth edition, Blackwell Publishing, ISBN 1-4051-3645-6. doi:10.1002/jctb.280440210
- Knipe, David M; Howley, Peter M; Griffin, Diane E; Lamb, Robert A; Martin, Malcolm A; Roizman, Bernard; Straus Stephen E. (2007) Fields Virology, Lippincott Williams & Wilkins. ISBN 0-7817-6060-7. doi:10.1046/j.1440-1711.2002.01083.x
- Shors, Teri (2008). Understanding Viruses. Jones and Bartlett Publishers. ISBN 0-7637-2932-9.
Liên kết ngoài
- Virus tại Encyclopædia Britannica (tiếng Anh)
- ViralZone - Nguồn của Viện Tin sinh học Thụy Sĩ về tất cả các họ virus, thông tin phân tử và dịch tễ học nói chung của chúng.
- Viruses — Open Access Journal – Tạp chí nghiên cứu mở MDPI
- PDBe Protein Data Bank in Europe Lưu trữ 2018-02-13 tại Wayback Machine Cấu trúc 3D của virus ở Ngân hàng Dữ liệu EM (EMDB)
(tiếng Việt)